

 science-review.ru
science-review.ru
Scientific journal
Scientific Review. Biological science
ISSN 2500-3399
ПИ №ФС77-57454
PECULIARITIES OF EMBRYONAL DEVELOPMENT OF THE COASTAL SWALL (RIPARIA RIPARIA L.) IN THE KAMA PRE-URAL (NORTH OF AREAL)

Проблемы размножения и раннего онтогенеза птиц в современной орнитологии остаются одними из актуальных. В публикациях, касающихся изучения биологии и экологии береговых ласточек, наиболее слабо исследован ранний онтогенез. Последние работы касаются в основном территориального размещения и численности колоний [1, 2]. В вышедших работах, посвященных экологии раннего онтогенеза диких птиц, материалы по ласточкам ограничены [3, 4]. Отсутствуют детальные исследования по темпу эмбрионального развития в период откладки яиц и на дальнейших этапах насиживания, о его вариабельности, величине разновозрастности, нет экспериментальных доказательств возможностей нормального развития зародышей при широком размахе уровней среднесуточных температур. Фрагментарными являются исследования, посвященные особенностям эмбриогенеза птенцовых, полуптенцовых и полувыводковых птиц, гнездящихся на границах ареалов [5].
Целью наших исследований было изучение особенностей эмбрионального развития береговой ласточки, размножающейся на севере ареала (Камское Предуралье).
Материалы и методы исследования
Материал собран в Соликамском районе (59,50 с.ш.) Пермского края в 2012 г. Темп яйцекладки у береговых ласточек изучали с помощью датированных кладок. Яйца, отложенные птицами в порядке снесения, метились быстросохнущей краской. Таким образом были получены датированные кладки. Анализ кладок проводили с учетом временного ранга яиц. Сутки инкубации отсчитывали со времени снесения последнего яйца. Этот метод позволил получить новые данные по темпу эмбрионального развития в период откладки яиц, о его вариабельности, величине разновозрастности зародышей на ранних этапах развития.
Сбор материала производили с периодичностью в 1–2 суток. Зародышей 1–12 стадий развития фиксировали в 10 %-ном растворе формалина с добавлением ледяной уксусной кислоты, затем окрашивали спиртовым борным кармином и заключали в глицерин-желатин. Стадии развития зародышей определяли по шкале, разработанной для воробьинообразных птиц.
При анализе степени развития эмбрионов первых – третьих суток инкубации учитывали величину первичной полоски, сегментацию осевой мезодермы, состояния головного мозга. Для сравнения величины разновозрастности в кладках, имеющих разное количество яиц, рассчитывали индекс разновозрастности как отношение разницы между крайними вариантами эмбрионов к числу яиц в кладке.
Результаты исследования и их обсуждение
В период размножения для ласточковых птиц характерен комплекс поведенческих реакций, связанных с заботой о потомстве: выбор места и постройка гнездового укрытия, охрана гнезда и кладки, регуляция факторов инкубации, выкармливание птенцов и ряд других. Важной адаптацией в раннем онтогенезе береговых ласточек является насиживание кладки с первого отложенного яйца, предотвращающее элиминацию зародышей.
Береговые ласточки относятся к группе птиц, насиживающих прерывисто, когда происходит постепенное увеличение плотности насиживания по мере накопления яиц в гнезде. Так же, в процессе яйцекладки у самок береговых ласточек последовательно осуществляется смена стадий (от первой до третьей) в развитии наседного пятна. Глубокие структурные изменения, происходящие в подкожной соединительной ткани, способствуют возрастанию теплоотдачи от тела самки к снесенным яйцам.
Таким образом, подогрев яиц во время периодических посещений семейными партнерами гнезда в ходе яйцекладки, формирование наседного пятна и постепенное увеличение плотности насиживания приводят к возникновению разновозрастности зародышей. Доказательством возникновения разновозрастности служат тотальные препараты зародышей из 0–1-суточных кладок (табл. 1, рисунок).
Таблица 1
Величина и индекс разновозрастности зародышей в 0- и 1-суточных кладках береговой ласточки
|
Вид |
Время завершения кладки, сутки |
Номер яйца стадия развития |
Величина разновозрастности |
Индекс разновозрастности |
|||||
|
1 |
2 |
3 |
4 |
5 |
в стадиях |
в часах |
|||
|
Береговая ласточка |
0 0 |
11 10 |
9 8 |
7 5 |
4 1 |
1 |
10 9 |
40–60 36–54 |
2,0 2,25 |
|
1 1 1 1 |
14 14 12 13 |
13 13 10 10 |
11 13 7 10 |
8 10 3 3 |
3 3 |
11 11 9 10 |
44–66 44–66 36–54 40–60 |
2,2 2,2 2,3 2,5 |
|
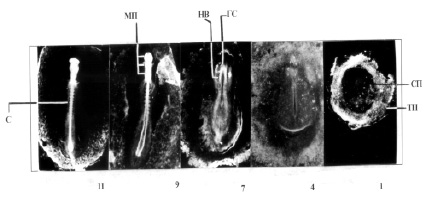
Разновозрастность зародышей береговой ласточки в 0-суточной полной кладке: ТП – темное поле, СП – светлое поле бластодермы, ГС – головная складка, НВ – нервные валики, МП – мозговые пузыри, С – сомиты; 11, 9, 7, 4, 1, – стадии развития
В 0-суточной пятияйцевой кладке береговой ласточки (рисунок) первый эмбрион находился на 11-й стадии развития, его длина была равна 8,2 мм. Величина сосудистого поля составляла 102,0 мм2. Видно, что первый мозговой пузырь у зародыша сжат в передне-заднем направлении. Сомитов 10 пар. Второй эмбрион соответствовал 9-й стадии развития. Его длина была равной 6,2 мм, величина сосудистого поля 63,6 мм2. Сомитов 7 пар. В головном отделе сформированы три первичных мозговых пузыря. Нервные валики в области сомитов сближены, в каудальном конце отстояли друг от друга на значительном расстоянии. Третий зародыш был на 7-й стадии. У него заметно начало сегментации осевой мезодермы, хорошо выделялись две пары сомитов. Длина зародыша равнялась 5,4 мм, величина сосудистого поля – 5,2 мм2. Четвертый эмбрион находился на 4-й стадии. Длина его составляла 3 мм. Светлое поле имело грушевидную форму. Было заметно образование на первичной полоске первичной бороздки. Последний зародыш был на первой стадии развития.
В односуточной кладке береговой ласточки, состоящей из пяти яиц, первый эмбрион находился на 14-й стадии. Длина зародыша составляла 10 мм, величина сосудистого поля – 132,6 мм2, в котором хорошо были видны формирующиеся кровяные островки. Количество сомитов – 18 пар, 7 из них прикрывал амнион. Угол между передним и задним мозгом был чуть больше прямого. Второй эмбрион несколько отставал в развитии от первого и находился на 13-й стадии. Количество сомитов – 16 пар, амнион прикрывал головную часть и 4 пары сомитов. Угол между передним и задним мозгом был тупым. Появились шейный и туловищный изгибы. Заметен поворот головы зародыша на левый бок, поэтому левый глазной бокал не виден. Менее сформирована и сосудистая система желточного мешка. Длина зародыша была равной 10 мм, величина сосудистого поля – 113,0 мм2. Третий эмбрион соответствовал 11-й стадии, его длина – 8,5 мм, величина сосудистого поля – 113,0 мм2. Сомитов 10 пар. Четвертый эмбрион находился на 8-й стадии развития. Длина его составляла 5,0 мм, величина сосудистого поля – 46,5 мм2. Сомитов 4 пары. Нервные складки сближены в области среднего и заднего мозга. Отчетливо видны границы сосудистого поля желточного мешка, в нем появились кровяные островки. Пятый, последний эмбрион кладки находился на 3-й стадии. Диаметр светлого поля был равен 4 мм, темного – 8 мм. Хорошо видна первичная полоска. Разновозрастность зародышей составила 11 стадий или 44–66 ч (1,8–2,8 суток). В других пятияйцевых кладках величина разновозрастности колебалась от 10 до 11 стадий, в четырехяйцевых от 9 до10, то есть от 36 до 60 ч (1,5–2,5 суток). Индекс разновозрастности варьировал от 2,0 до 2,5 единицы.
Таким образом, у береговых ласточек в завершенных кладках, состоящих из пяти яиц, величина разновозрастности эмбрионов варьировала от 10 до 11 стадий, в четырехъяйцевых кладках колебалась от 9 до 10 стадий. По времени данный показатель составил от 36 до 66 ч, или от 1,5 до 2,8 суток в разных кладках. Величина индекса разновозрастности варьировала от 2,0 до 2,5 единицы. Разное время пребывания семейных партнеров на гнезде является показателем индивидуальной изменчивости процессов насиживания у каждой гнездящейся пары. Большая вариативность в начальный период насиживания у пар береговушек связана, очевидно, с прерывистой инкубацией яиц во время яйцекладки.
Дальнейший анализ таблицы показывает, что чем раньше было отложено яйцо, тем более развитым был в нем зародыш. Более близкими по степени развития являются эмбрионы из первых-третьих яиц (отличие в 1–2 стадии). Различия в развитии между последними и предпоследними эмбрионами достигали 3–4 стадий, в одном случае даже 7. Следует отметить, что в разных кладках различия между эмбрионами с учетом временного ранга неодинаковы и колеблются в пределах 1–7 стадий, чаще 1–4. Это еще раз подтверждает индивидуальный характер насиживания в разных кладках.
Наши исследования по изучению темпа эмбриогенеза береговых ласточек свидетельствуют о том, что начиная со времени яйцекладки зародыши развиваются гетерохронно. Так, в односуточных кладках из пяти яиц первые эмбрионы достигли 12– 14-й стадии, а яйца находились в гнездах уже 6 суток и при максимальном темпе эмбрионального развития, под которым понимают развитие в последнем или предпоследнем яйце, они могли бы достичь 25–26-й стадии. Вторые и третьи эмбрионы так же были развиты меньше ожидаемого. Следовательно, зародыши из первых яиц развиваются в замедленном темпе. Зародыши из четвертых, пятых яиц развиваются в максимальном темпе, и поэтому видна прямая зависимость между временем с момента откладки и сроком инкубации. Разный темп развития эмбрионов можно объяснить поведением самки на гнезде в период яйцекладки и степенью сформированности у нее наседного пятна. В начальный период яйцекладки плотность насиживания низкая, а наседное пятно характеризуется первой стадией развития. К завершению откладки яиц плотность насиживания у береговушек приближена к максимальной, а развитие наседного пятна достигает третьей стадии. Таким образом, яйца, отложенные последними, попадают под воздействие более высоких температур и эмбрионы в них развиваются с оптимальной скоростью.
В период собственно насиживания происходит дальнейшее увеличение плотности насиживания, наседное пятно проходит в своем развитии от 3-й до 5-й стадии, нарастает внутригнездовая температура. Поэтому разновозрастность зародышей, возникшая у береговых ласточек в период откладки яиц, сохраняется и в период собственно насиживания.
Так, в пятияйцевой кладке береговой ласточки, взятой на анализ через трое суток инкубации (со времени снесения последнего яйца) первый эмбрион по степени развития соответствовал 21-й стадии, второй – 18-й стадии, третий –14-й стадии, четвертый – 13-й, а пятый – на 12-й стадии развития. Величина разновозрастности составила 9 стадий.
На четвертые сутки собственно насиживания первый зародыш из трехъяйцевой кладки находился на 24-й стадии, второй зародыш на 22-й стадии, а третий 19-й стадии развития. Разница между первым и третьим эмбрионами была равной 5 стадиям.
На пятые сутки инкубации зародыши из трехяйцевой кладки соответствовали 27-й, 25-й и 23-ей стадиям развития. Величина разновозрастности составила 4 стадии.
На шестые сутки инкубации самый старший зародыш соответствовал 31-й стадии в развитии, второй – 29-й стадии, третий эмбрион был на 28-й стадии, четвертый эмбрион находился на 26-й стадии. Величина разновозрастности между крайними вариантами составила 5 стадий.
На седьмые сутки инкубации в четырехъяйцевой кладке первый эмбрион находился на 32-й стадии развития, второй на 31-й стадии, третий – на 29-й стадии, четвертый – на 28-й стадии. Таким образом, последний зародыш отставал в развитии от первого на 4 стадии.
После девяти суток инкубации зародыши из трехяйцевой кладки были соответственно на 36-й, 35-й, 33-ей стадиях. Разновозрастность между первым и третьим эмбрионами равнялась трем стадиям.
На 10-е сутки собственно насиживания первый эмбрион в кладке из четырех яиц находился на 37-й стадии развития, второй зародыш был на 36-й стадии, третий – на 35-й. Четвертый соответствовал 34-й стадии. Разница между крайними вариантами в кладке составила 3 стадии.
На 12 сутки собственно насиживания в кладке из четырех яиц первый эмбрион находился на 42-й стадии, второй на 39-й стадии, а третий и четвертый эмбрионы соответствовали 38-й стадии. Разновозрастность между первым и четвертым эмбрионами равнялась четырем стадиям.
В 13-суточной трехъяйцевой кладке первый эмбрион находился на 42-й стадии. Второй зародыш был на 41-й стадии. Третий зародыш соответствовал 40-й стадии. Разница между крайними вариантами в кладке составила 2 стадии.
На 14-е сутки после завершения кладки в некоторых гнездах началось вылупление птенцов. Птенцы береговых ласточек появлялись на свет в редком эмбриональном пуху, развитом на следующих птерилиях: головной (по 3–5 перьевых зачатков над каждым глазом длиной 6–7 мм), затылочной (6 зачатков длиной 6–7 мм), плечевой (по 2–5 зачатков длиной 7,5–8,5 мм), спинной (по 4–5 зачатков длиной 7,5–10,5 мм). Эмбриональный пух распушался спустя 2–3 ч после вылупления птенца из яйца. Длина крыла варьировала от 10 до 11 мм. Цевка имела длину 4 мм, третий палец ноги – 4 мм. Длина клюва от угла рта до его кончика была равна 3,5–6,0 мм от ноздри до кончика 1,0–1,2 мм. Цвет клюва – белый. Количество щитков на цевке равнялось 5, на третьем пальце ноги – 10. Форма когтей – крючкообразная. Масса вылупившихся птенцов в среднем составила 1,1 ± 0,08 г (n = 8, lim = 1,0–1,2 г), то есть 79–82 % от массы свежеснесенного яйца. Порядок вылупления птенцов соответствовал порядку откладки яиц. Растянутость вылупления птенцов в отдельных кладках колебалась от 20 до 24 ч.
Величина и индекс разновозрастности эмбрионов в кладках береговых ласточек представлены в табл. 2.
Таблица 2
Величина и индекс разновозрастности эмбрионов в кладках береговой ласточки при естественной инкубации
|
Показатели |
Сутки инкубации |
||||||||||
|
1 |
3 |
4 |
5 |
6 |
7 |
9 |
10 |
12 |
13 |
14 |
|
|
Стадия развития зародыша |
|||||||||||
|
из первого яйца |
14 |
21 |
24 |
27 |
31 |
32 |
36 |
37 |
40 |
42 |
птенец |
|
из последнего яйца |
3 |
12 |
19 |
23 |
26 |
28 |
33 |
34 |
38 |
40 |
43 |
|
Величина разновозрастности в стадиях |
11 |
9 |
5 |
4 |
5 |
4 |
3 |
3 |
3 |
2 |
|
|
Индекс разновозрастности |
2,2 |
1,8 |
1,7 |
1,3 |
1,25 |
1,0 |
1,0 |
0,75 |
0,75 |
0,6 |
|
|
Количество яиц в кладке |
5 |
5 |
3 |
3 |
4 |
4 |
3 |
4 |
4 |
3 |
|
Из табл. 2 следует, что до третьих суток инкубации (со времени снесения последнего яйца) величина разновозрастности составляет 9–11 стадий, далее с 4-х по 12-е сутки собственно насиживания разница между крайними вариантами не превышает 3–5 стадий. Величина индекса разновозрастности, начиная с первых суток инкубации, постепенно уменьшалась с 2,2 до 0,6 единицы.
Следствием уменьшения величины разновозрастности в кладках береговых ласточек к концу эмбрионального развития является дружное вылупление птенцов: по времени за меньший срок, чем можно было бы ожидать на основании величины разновозрастности эмбрионов при завершении откладки яиц.
Разница в темпе эмбрионального развития зародышей из первых и последних яиц кладки несколько сглаживается в конце эмбриогенеза за счет проявления компенсаторного роста и создает условия для более дружного вылупления птенцов и лучшего выживания в постнатальном онтогенезе.
Выводы
Для береговых ласточек, гнездящихся на севере ареала, характерно постепенное увеличение плотности насиживания, результатом которого является гетерохронность развития эмбрионов. Величина разновозрастности зародышей подвержена значительным колебаниям и по времени может составлять от 1,0 до 2,5 суток, что свидетельствует о широкой индивидуальной изменчивости процессов насиживания каждой пары. Темп развития эмбрионов к концу яйцекладки в пределах одной отдельно взятой кладки неодинаков, он является замедленным для зародышей из яиц низших временных рангов, что связано с поведением наседки (семейных партнеров) на гнезде и структурными изменениями, происходящими в развитии наседного пятна. В период собственно насиживания яйца, отложенные последними, попадают в более высокие температурные условия и зародыши в них развиваются с оптимальной скоростью. Разный темп развития эмбрионов из яиц высшего и низшего временных рангов обеспечивает не только сохранение жизнеспособности в период яйцекладки, но и приводит к меньшей растянутости вылупления птенцов (в сравнении с ожидаемой), а значит, и большей вероятности их выживания в постнатальном онтогенезе.
Библиографическая ссылка
Сугробова Н.Ю. ОСОБЕННОСТИ ЭМБРИОНАЛЬНОГО РАЗВИТИЯ БЕРЕГОВОЙ ЛАСТОЧКИ (RIPARIA RIPARIA L.) В КАМСКОМ ПРЕДУРАЛЬЕ (СЕВЕР АРЕАЛА) // Научное обозрение. Биологические науки. 2018. № 6. С. 25-30;URL: https://science-biology.ru/en/article/view?id=1125 (дата обращения: 26.07.2026).