

 science-review.ru
science-review.ru
Scientific journal
Scientific Review. Biological science
ISSN 2500-3399
ПИ №ФС77-57454
THE DENSITY OF MICROCENOSIS PINE AND ITS EFFECT ON THE SIZE OF TREES AND GROUND COVER

В структуре древостоев микроценозы играют важную роль во внутри- и межвидовых взаимодействиях между деревьями и растениями и для их изучения применяют целый спектр разнообразных методов пространственного анализа и связывают площадь питания дерева с диаметрами деревьев и диаметром крон [1–3], включая высоту дерева и протяженность кроны [4], а в наиболее сложных моделях конкурентного взаимовлияния деревьев также с массой хвои и текущим приростом [5]. В отдельных случаях определяют динамику прироста с еженедельными измерениями у одиночных деревьев и деревьев в биогруппах [6].
В настоящей работе использован наиболее простой метод выделения микроценозов с механическим делением территории на площадки размером 10×10 м. Такие «механические» микроценозы лишены внешней физиономичности и выхвачены из структуры связей с соседями [1], однако при таком их выделении полностью исключен субъективный фактор, что немаловажно. При больших выборках вполне можно получить результаты с достаточно точными статистическими оценками без влияния тех или иных предпочтений исследователя. А такие предпочтения неизбежны при выборе мест выделения микроценозов, когда их выбирают, на взгляд исследователя, в неких «типичных» местах, например вдали от окон и прогалин [7]. Такие методы дают смещение в оценках влияния структур фитоценоза на размерные характеристики дерева в сторону их увеличения, так как окна, а также скопления деревьев в виде биогрупп являются неотъемлемым свойством древостоя и исключать их при выборе модельных площадок некорректно [8].
При механическом методе выделения микроценозов таких нежелательных предпочтений можно избежать, используя, например, для ботанического описания напочвенного покрова мини-площадки размером 2×2 м, которые закладывают в центрах внутри площадок 10×10 м, что и было применено в данной работе.
Цель данной работы: выяснить количественные значения силы влияния фактора густоты микроценозов сосны на размеры деревьев, а также на число и частоту встречаемости видов растений напочвенного покрова.
Материалы и методы иследования
184-летнее насаждение сосны 1 класса бонитета на площади 2,16 га с полнотой 0,93, тип леса сосняк чернично-кисличниковый. У деревьев делали отметку на высоте 1,3 м, наносили номер, измеряли окружность ствола и определяли диаметр с округлением до ± 0,1 см. Деревья наносили на план с точностью ±10–35 см, что потребовало несколько месяцев полевых и камеральных работ. Всего было измерено 735 деревьев. План был оцифрован в программе ArcMap-ArcViev и механически разбит на 216 ячеек (микроценозов) размером 10×10 м. При ботаническом описании растительности использовали методы полевых экологических исследований [9, 10] с использованием коротких транссект длиной до 180 м, которые названы здесь «треками». Всего было заложено 13 треков, из них в данном массиве сосны проложены два под номерами 3 и 4. На каждом строго через 10 м и в центре площадок 10×10 м закладывали микроплощадки 2×2 м, где проводили описание растительности. Была принята следующая градуировка частот встречаемости растений в баллах: 1 – единично, 2 – от 5 до 10 шт., 3 – 11–30 шт., 4 – 31–50 шт., 5 – более 50 шт. Для мхов глазомерно определяли покрытие поверхности: 1 – до 10 %; 2 – 11–25 %; 3 – 26–50 %; 4 – 51–80 %; 5 – более 80 %.
Результаты исследования и их обсуждение
Расположение треков 3 и 4 в насаждении показано ниже (рис. 1).

Рис. 1. Площадки 10×10 м с тонировкой по числу деревьев, обозначенных точками. Площадь территории 2,16 га. На треке 4 заложено 17 миниплощадок 2×2 м для ботанического описания напочвенного покрова в ячейках от № 0 до № 204 (с пропуском ячейки 132). На треке 3 заложено 16 миниплощадок 2×2 м в ячейках от № 19 до № 199
Всего в средней и западной части массива сосны на треках 3 и 4 заложено 33 миниплощадки размером 2×2 м. Расположение треков в пределах изучаемого участка показано на плане, где густота микроценозов сосны была градуирована на 8 позиций – от отсутствия деревьев (нет окраски ячейки) и до 7 шт. деревьев в ячейках с самой темной окраской (рис. 1).
Анализ результатов показал следующее. Плотность стояния деревьев в микроценозе, которую измеряли числом деревьев на площадках 10×10 м, повлияла на диаметр сосны с силой 9,4 %, если оценивать ее по показателю аппроксимации (R2 = 0,094) для тренда полинома третьей степени (рис. 2).
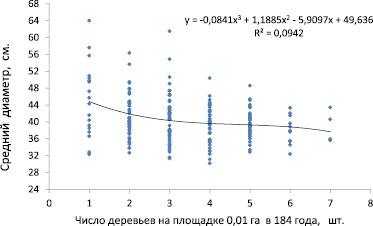
Рис. 2. Влияние густоты стояния деревьев сосны на площадках 10×10 м на средний диаметр деревьев в 184 года: прерывистая линия – средний диаметр деревьев древостоя
Если же взять линейный тренд для этой выборки из 199 площадок (7 площадок не имели деревьев), то он имеет показатель R2 = 0,078 и достоверную корреляцию r = –0,30 ± 0,06. Дополнительно были построены полиномы 2–6 степени, и для каждого из них определили показатели аппроксимации R2, из которых рассчитали среднее значение. В результате расчетов по этим шести показателям силы влияния (с учетом коэффициента детерминации, рассчитанного по квадрату корреляции) оказалось, что число деревьев на площадке 10×10 м (т.е. густота микроценоза) повлияло на средний диаметр сосны на них в возрасте 184 года с силой в среднем 8,8 %.
Эти данные характеризуют площадки 0,01 га как микроценозы с очень слабым (8,8 %) итоговым влиянием их густоты на размер входящих в них деревьев. При таком малом влиянии локальной густоты можно считать, что конкуренция между деревьями в микроценозах уже не является тем фактором, который в решающей степени определял бы размерные характеристики деревьев. Это противоречит парадигме о сильном влиянии густоты на размер деревьев в насаждении.
Однако в микроценозах все оказалось не так просто, и влияние их локальной густоты оказалось резко ослаблено. В литературе, посвященной изучению конкуренции между деревьями, этому явлению есть подтверждение [5, 11]. Для нашего насаждения сосны ранее удалось выяснить, что при анализе биогрупп рост сосны в них оказался одинаковым со свободно растущими деревьями, несмотря на громадные различия в расстояниях до соседей, а диаметры деревьев в них коррелировали с r = 0,47–0,58, что позволило отнести взаимодействие деревьев в биогруппах к партнерскому типу [12], которое в несколько раз сильнее давления конкуренции и, по-видимому, замещает ее в спелом возрасте.
Следующей задачей было изучение влияния густоты площадок размером 0,01 га, отражающих локальную сомкнутость верхнего яруса насаждения, на число видов растений в напочвенном покрове и на частоту их встречаемости на мини-площадках размером 2×2 м. Влияние на обилие видов изучали по двум показателям заполнения пространства деревьями: первый по числу деревьев на площадке 0,01 га и второй по сумме площадей сечения стволов на высоте 1,3 м, которое в среднем по массиву сосны составило 0,442 м2 на 1 площадку. Для диаграмм сечения были выражены в процентах от последней величины (рис. 3).
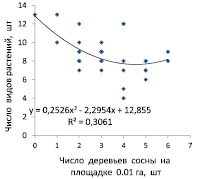
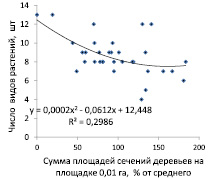
Рис. 3. Влияние показателей заполнения пространства деревьями сосны на площадке 0,01 га: по числу деревьев (слева) и по сумме площадей сечения стволов (справа) на число видов растений в напочвенном покрове
Оказалось, что по показателям аппроксимаций трендов R2 этих диаграмм, равных 0,306 и 0,298, число видов растений в напочвенном покрове обусловлено густотой верхнего яруса сосны на 30,6 %, а таксационная полнота, выраженная в виде суммы площадей сечения деревьев, влияет с силой 29,8 %. Эти оценки показывают, что сила влияния плотности верхнего яруса древостоя на число видов растений напочвенного покрова равна в среднем 30,2 %. По своему уровню это средняя сила влияния, при которой наибольшее число видов растений (7–13 шт.) встречается при снижении густоты верхнего яруса до 0–3 шт. деревьев на площадке 0,01 га, что оказывается ниже средней густоты.
Далее была рассчитана средняя частота встречаемости растений и проективное покрытие моховым покровом на треках 3 и 4 (рис. 4).
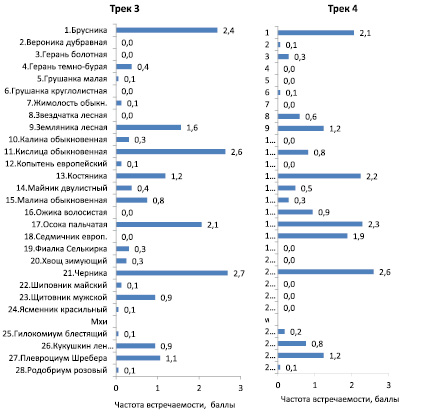
Рис. 4. Средняя частота встречаемости растений в баллах для видов растений напочвенного покрова на треках 3 и 4 в сосняке чернично-кисличниковом
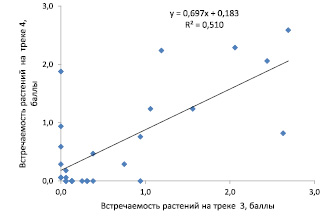
Рис. 5. Связь между средней частотой встречаемости растений для 28 видов растений напочвенного покрова на треках 3 и 4 в сосняке чернично-кисличниковом
Доминирующими видами на обоих треках являются 6 видов со средней частотой встречаемости более 1 балла: брусника, земляника лесная, костяника, осока пальчатая, черника, плевроциум Шребера. Для оценки сходства частот встречаемости видов на треках можно использовать их корреляцию, размещая частоты для трека 3 по горизонтальной, а для трека 4 – по вертикальной оси точечной диаграммы (рис. 5).
Полученное на диаграмме облако точек показывает, что если растения встречаются на одном треке с частотой выше 1 балла (от 11 шт. растений и более), то на другом треке их частота также оказывается достаточно высокой. Однако встретились и исключения: седмичник европейский (№ 18, рис. 4) на треке 3 не встречался, тогда как на треке 4 его встречаемость оказалась на уровне 1,9 балла (рис. 4).
Для частот встречаемости растений на треках 3 и 4 их взаимная корреляция оказалась достаточно сильной и равной r = 0,71 ± 0,09. Показатель достоверности аппроксимации линейного тренда R2 = 0,510 показывает, что частота встречаемости видов на одном треке обуславливает их частоту на другом на 51 %, что можно считать высоким значением.
Выводы
1. Расчеты по шести показателям силы влияния, включая коэффициент детерминации по квадрату корреляции, равной r = –0,30 ± 0,06, показали, что число деревьев сосны на площадке 10×10 м повлияло на их средний диаметр в возрасте 184 года с силой в среднем всего лишь 8,8 %. Это противоречит парадигме, по которой влияние густоты на уровне древостоя в целом должно быть сильным. Однако на уровне микроценоза влияние локальной густоты оказалось резко ослаблено до малозначимых значений силы ее влияния.
2. Наибольшее число видов растений в напочвенном покрове (7–13 шт.) встречается при снижении густоты яруса сосны до 0–3 шт. деревьев на площадках размером 10×10 м, т.е. ниже средней густоты; при этом сила влияния густоты и таксационной полноты яруса сосны на число видов растений составляет в среднем 30,2 %.
3. Частота встречаемости каждого из видов растений на одном треке обуславливает частоты их встречаемости на другом треке на 51 %, что позволяет считать части насаждения, где они заложены, достаточно однородными по видовому составу растений.
Библиографическая ссылка
Рогозин М.В., Меркулов А.Д. ГУСТОТА МИКРОЦЕНОЗОВ СОСНЫ И ЕЕ ВЛИЯНИЕ НА РАЗМЕР ДЕРЕВЬЕВ И НАПОЧВЕННЫЙ ПОКРОВ // Научное обозрение. Биологические науки. 2019. № 2. С. 41-46;URL: https://science-biology.ru/en/article/view?id=1145 (дата обращения: 24.06.2026).