

 science-review.ru
science-review.ru
Scientific journal
Scientific Review. Biological science
ISSN 2500-3399
ПИ №ФС77-57454
THYROID STATUS OF THE ORGANISM IN THE INDUCTION AND INHIBITION OF THE MONOOXYGENASE LIVER SYSTEM IN EXPERIMENT

Гормоны щитовидной железы – тироксин (Т4) и трийодтиронин (Т3) являются необходимыми регуляторными молекулами для развития, роста и функционирования клеток и органов. Они регулируют уровень базального метаболизма клеток, в том числе и гепатоцитов. Следовательно, изменение уровня тиреоидных гормонов должно сказываться на функционировании печени. В то же время в печени с помощью ферментов-дейодиназ протекает дейодирование тиреоидных гормонов. Следовательно, функциональная активность гепатоцитов также может повлиять на содержание тиреоидных гормонов. Известно, что Т3 имеет большее сродство к ядерным рецепторам, чем Т4. В то же время Т4 секретируется в 10 раз больше, чем Т3. Дейодирование Т4 до Т3 идёт с помощью трех групп ферментов-дейодиназ: D1, D2 и D3. Эти ферменты отвечают как за превращение Т4 в более активный Т3, так и инактивацию Т4 путём превращения в обратный Т3 и превращение обратного Т3 и Т3 в дийодтиронин. Превращение Т4 в Т3 в тканях происходит под действием дейодиназ типа D1, которая обнаруживается главным образом в печени и почках. Основную роль в инактивацию как Т4, так и Т3 играет дейодиназа D3. Она локализована в печени, коже и центральной нервной системе. Кроме участия в метаболизме в печени синтезируется ряд плазматических белков, связывающих тиреоидные гормоны: тироксинсвязывающий глобулин, тироксинсвязывающий преальбумин и альбумин. При большинстве хронических заболеваний наблюдаются нарушения метаболизма тиреоидных гормонов, характеризующиеся нормальным уровнем общего Т4, нормальным или повышенным уровнем свободного Т4 (сТ4), низким уровнем общего Т3, низким уровнем свободного Т3 (сТ3) и повышенным уровнем обратного Т3. Это состояние называется синдромом эутиреоидной патологии (sick euthyroid syndrome) [1]. Выявление зависимости между тиреоидным статусом организма и функционированием печени позволит в дальнейшем разработать методы, позволяющие регулировать функционирование как самой печени, так и щитовидной железы.
Цель исследования: оценка зависимости тиреоидного статуса организма от активности микросомальной монооксигеназной системы печени в эксперименте у крыс.
Материалы и методы исследования
Опыты проведены на 70 белых крысах-самцах массой 180–220 г. Животные содержались в стандартных условиях вивария при естественном освещении и свободном доступе к воде и пище. Протокол экспериментов соответствовал этическим нормам, изложенным в «Правилах проведения работ с использованием экспериментальных животных», а также в Директиве 2010/63/EU Европейского парламента и Совета Европейского союза по охране животных, используемых в научных целях.
Индукцию монооксигеназной системы печени у 40 крыс вызывали её индукторами – бензоналом (бензобарбитал) и зиксорином (флумецинол). У 15 крыс проводили индукцию монооксигеназной системы печени пероральным введением бензонала на крахмальном клейстере в течение трех дней в дозе 50 мг/кг веса. У 15 крыс проводили индукцию монооксигеназной системы печени пероральным введением зиксорина на крахмальном клейстере в течение четырех дней в дозе 40 мг/кг веса. Ингибирование монооксигеназной системы печени у 40 крыс вызывали её ингибиторами – хлорида кобальта (CoCl2) и циметидином. У 15 крыс проводили ингибирование монооксигеназную систему печени одноразовым внутрибрюшинным введением CoCl2 в дозе 30 мг/кг веса. У 15 крыс проводили ингибирование монооксигеназной системы печени пероральным введением циметидина на крахмальном клейстере в течение 10 дней в дозе 100 мг/кг веса. 10 крыс служили контролем.
Для оценки метаболической активности монооксигеназной системы печени проводили гексеналовый тест. Гексенал животным вводили в дозе 100 мг/кг, внутрибрюшинно. Учитывалось время между потерей и приобретением «рефлекса переворачивания». При этом животных помещали в термостатирующую камеру с температурой 26 °С. После пробуждения крысы забивались под эфирным наркозом путём декапитации и собиралась кровь. В сыворотке крови определяли содержание свободного и общего трийодтиронина и тироксина, тиреотропного гормона (ТТГ) методом твердофазного иммуноферментного анализа ELISA, при помощи тест-систем фирмы «Human» (Германия) на микропланшетном фотометре MR96A (Mindray, Китай).
Печень крыс гомогенизировали в стеклянном гомогенизаторе с тефлоновым пестиком в среде выделения, состоящей из 0,25 М сахарозы, 0,05 М KCl в растворе 0,05 М трис-HCl буфера, рН = 7,4. Гомогенат подвергался дифференциальному центрифугированию и в микросомальной фракции определяли содержание и активность компонентов монооксигеназной системы. Содержание цитохрома Р-450 в микросомальной суспензии определяли по методу T. Omura, R. Sato (1964). Содержание цитохрома b5 определяли после восстановления опытных образцов суспензии микросом при добавлении НАДН. Скорость п-гидроксилирования анилина в микросомальной фракции оценивали по образованию п-аминофенола, а N-деметилирования амидопирина в микросомально-цитозольной фракции по образованию формальдегида. Содержание белка в пробах определяли по Lowry et.al.
Полученные цифровые результаты были обработаны с помощью стандартных методов вариационной статистики с применением t-критерия Стьюдента.
Результаты исследования и их обсуждение
Для выявления зависимости между тиреоидным статусом организма и функционально активным состоянием монооксигеназной системы печени в качестве индукторов данной системы были выбраны два индуктора – бензонал и зиксорин.
Бензонал (1-бензоил-5-этил-5-фенил- барбитуровая кислота) является индуктором цитохрома Р-450 фенобарбиталового типа. Под его влиянием в печени существенно увеличивается содержание микросомального белка, цитохрома Р-450 и активность НАДФН цитохром Р-450 редуктазы. Действительно, результаты исследований показали, что у животных с бензоналовой индукцией монооксигеназной системы печени длительность гексеналового сна оказалась укороченной на 36,6 % по сравнению с интактным показателем (таблица).
Содержание и активность компонентов микросомальной монооксигеназной системы печени при индукции и ингибировании
|
Длительность гексеналового сна, мин |
Содержание микросомальных цитохромов, нмоль/мг белка |
Активность микросомальных ферментов, нмоль/мин • мг белка |
|||
|
Р-450 |
b5 |
Анилингид-роксилаза |
Амидопирин-N-деметилаза |
||
|
Контроль |
28,00 ± 0,87 |
0,99 ± 0,09 |
0,41 ± 0,03 |
0,94 ± 0,08 |
2,79 ± 0,26 |
|
Бензонал |
17,75 ± 0,75* |
1,52 ± 0,13* |
0,48 ± 0,03 |
1,29 ± 0,11* |
4,22 ± 0,41* |
|
Зиксорин |
22,13 ± 2,5* |
1,39 ± 0,08* |
0,50 ± 0,05 |
1,19 ± 0,04* |
5,02 ± 0,48* |
|
CoCl2 |
48,63 ± 0,25* |
0,44 ± 0,03* |
0,28 ± 0,04* |
0,53 ± 0,08* |
1,22 ± 0,07* |
|
Циметидин |
51,25 ± 0,25* |
0,40 ± 0,06* |
0,25 ± 0,03* |
0,39 ± 0,04* |
1,02 ± 0,08* |
Примечание: * – Р < 0,05 по сравнению с интактными показателями.
При этом содержание основного компонента монооксигеназной системы – цитохрома Р-450 было повышено на 53,5 % от интактного значения. Хотя абсолютное значение цитохрома b5 было повышено от интактного значения на 17,1 %, однако данное различие от контроля оказалось статистически не значимым (Р > 0,05). Анилингидроксилазная и амидопирин-N-деметилазная активность микросом при бензоналовой индукции оказалась выше интактных значений на 37,2 и 51,3 % соответственно.
Зиксорин по химической структуре является 3-фторметил-α-этилбензгидролом. Он также увеличивает содержание компонентов микросомальной монооксигеназной системы печени. Под его влиянием усиливается образование глюкуронидов и выделение желчи. У животных с индукцией монооксигеназной системы печени зиксорином длительность гексеналового сна была укорочена на 21,0 % по сравнению с интактной величиной (таблица).
Зиксориновая индукция также привела к увеличению содержания цитохрома Р-450. Так, оно оказалось выше интактного значения на 40,4 %. В содержании цитохрома b5, аналогично бензоналовой индукции, хотя наблюдалось повышение на 22,0 % от контроля, однако и это повышение оказалось статистически незначимым. При зиксориновой индукции анилингидроксилазная активность микросом оказалась выше интактных значений на 21,4 %, а амидопирин-N-деметилазная – на 79,9 %.
Следовательно, бензонал и зиксорин приводят к значительной индукции монооксигеназной системы печени.
Для выявления зависимости между тиреоидным статусом организма и функционально угнетённым состоянием монооксигеназной системы печени в качестве ингибиторов данной системы были выбраны два вещества – CoCl2 и циметидин.
Результаты исследований показали, что у животных с ингибированием монооксигеназной системы печени CoCl2 длительность гексеналового сна оказалась удлинённой на 73,7 % по сравнению с интактным показателем (таблица). При этом содержание основного компонента монооксигеназной системы – цитохрома Р-450 было снижено на 55,6 % от интактного значения. Содержание цитохрома b5 было снижено от интактного значения на 31,7 %. Анилингидроксилазная и амидопирин-N-деметилазная активность микросом при ингибировании монооксигеназной системы оказалась ниже интактных значений на 43,6 и 56,3 % соответственно.
При введении циметидина экспериментальным животным длительность гексеналового сна была удлинена на 83,0 % по сравнению с интактной величиной (таблица). Циметидиновая ингибиция также привела к уменьшению содержания цитохрома Р-450. Так, оно оказалось ниже интактного значения на 59,6 %. В содержании цитохрома b5 наблюдалось снижение на 39,0 % от интактного значения. При циметидиновой ингибиции монооксигеназной системы анилингидроксилазная активность микросом оказалась ниже интактных значений на 58,5 %, а амидопирин-N-деметилазная – на 63,4 %.
Следовательно, хлорид кобальта и циметидин приводят к значительному ингибированию монооксигеназной системы печени.
Изучение тиреоидного статуса при бензоналовой индукции монооксигеназной системы печени показало статистически значимое увеличение в крови содержания общего Т3 на 23 % и свободного – на 11,7 % от интактных значений (рис. 1).
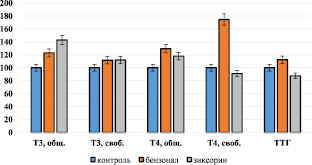
Рис. 1. Тиреоидный статус организма при бензоналовой и зиксориновой индукции монооксигеназной системы печени. По оси ординат содержание показателей в %
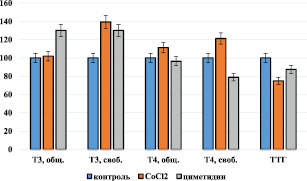
Рис. 2. Тиреоидный статус организма при ингибировании монооксигеназной системы печени кобальтом хлорида и циметидином. По оси ординат содержание показателей в %
В содержании Т4 также наблюдали повышение. Так, содержание общего Т4 было увеличено на 29,4 %, а свободного – на 74,6 % от контроля. Абсолютное значение ТТГ, хотя было увеличено на 12,5 % по сравнению с контролем, однако статистически было незначимо.
Изучение тиреоидного статуса при зиксориновой индукции монооксигеназной системы печени также показало статистически значимое увеличение в крови содержания общего Т3 на 43 % и свободного – на 12,0 % от интактных значений (рис. 1). Содержание общего Т4 было увеличено на 18,0 %, а в содержаниях свободного Т4 и ТТГ статистически значимых изменений от интактных показателей не выявлено.
Таким образом, полученные результаты показали, что при индукции монооксигеназной системы печени её индукторами – бензоналом и зиксорином – наблюдается увеличение содержания гормонов щитовидной железы – Т3 и Т4 в крови, на фоне практического неизменения уровня ТТГ.
Изучение тиреоидного статуса при ингибировании монооксигеназную систему печени хлоридом кобальта показало статистически значимое увеличение в крови содержания свободного Т3 на 39,3 % от интактного значения, на фоне неизменения его общего содержания (рис. 2).
Увеличение содержания общего Т4 на 11,3 % от интактного значения оказалось статистически незначимым (Р > 0,05), однако содержание свободного Т4 оказалось выше контроля на 21,3 %. В содержании ТТГ при ингибировании монооксигеназной системы хлоридом кобальта наблюдали статистически значимое снижение от контроля на 25,0 %.
Изучение тиреоидного статуса при циметидиновом ингибировании монооксигеназной системы печени показало статистически значимое увеличение в крови как содержания общего, так и свободного Т3 на 30,2 и 30,1 % соответственно от интактных значений (рис. 2). В содержании общего Т4 статистически значимое изменение от интактного значения не наблюдали. В то же время содержание свободного Т4 было снижено на 21,1 %. Содержание ТТГ оказалось статистически значимо сниженным на 12,5 % по сравнению с контролем.
Таким образом, полученные результаты показали, что при ингибировании монооксигеназной системы печени её ингибиторами – хлоридом кобальта и циметидином – наблюдается некоторое увеличение содержания гормонов щитовидной железы – Т3 и Т4 в крови, на фоне снижения содержания ТТГ.
В исследовании для индукции в монооксигеназную систему печени были использованы «эталонные» индукторы микросомального окисления – бензонал и зиксорин. Хотя оба они относятся к фенобарбиталовому типу индукторов [2], однако различаются по «спектру» индуцируемых изоформ цитохрома P-450. Бензонал индуцирует изоформы цитохрома P-450IIB, P-450IIC, P-4501IIA, а зиксорин – изоформы цитохрома Р-450IA, P-450IIB. Введение бензонала крысам вызывает увеличение содержания цитохрома Р-450 и активность НАДФН цитохром Р-450 редуктазы, на фоне быстрой активации транскрипции CYP2B1 и CYP2B2 [3]. Под влиянием бензонала довольно сильно усиливается также синтез белка [4], что подтверждается также морфологическими исследованиями. Так, бензоналовая индукция, аналогично фенобарбиталовой, проявляется увеличением объёма гепатоцитов за счёт увеличения объёма их цитоплазмы и в меньшей степени ядер. Так, М.В. Захаровой показано, что фенобарбиталовая индукция приводит к увеличению объёма гепатоцитов, их цитоплазмы и ядер соответственно на 74, 77 и 42,7 % от контроля [5]. В то же время в данной работе показано, что при зиксориновой индукции объём гепатоцитов, цитоплазмы и ядер увеличивался соответственно на 32, 33 и 27,3 % по сравнению с контролем. При введении зиксорина, в отличие от бензонала, не наблюдается усиление синтеза белка [4].
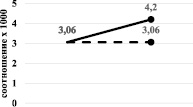
Вероятно, именно это и явилось причиной разнонаправленного изменения содержания тиреоидных гормонов при бензоналовой и зиксориновой индукции. Для наглядности этих изменений мы проанализировали изменения при различной индукции монооксигеназной системы печени соотношения сТ3/Т3 и сТ4/Т4. Результаты показали, что при бензоналовой индукции наблюдается снижение соотношения сТ3/Т3 и повышение сТ4/Т4 (рис. 3, а), а при зиксориновой индукции снижение как сТ3/Т3, так и сТ4/Т4 (рис. 3, б).
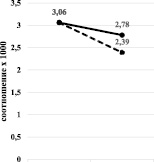
а) б)
Рис. 3. Изменение соотношения сТ3/Т3 (а) и сТ4/Т4 (б) при бензоналовой (сплошная линия) и зиксориновой (прерывистая линия) индукции
Одним из возможным механизмов данных изменений может быть именно специфическое влияние использованных индукторов на скорость синтеза белка. Бензоналовая индукция сильно индуцирует также синтез белка. T3 и T4 в крови циркулируют в связанными с тироксинсвязывающим глобулином, тироксинсвязывающим преальбумином и альбумином. В то же время именно свободный T3 и T4 влияют на все этапы метаболизма, роста и развития, осуществляя поддержание температуры тела и стимулируя теплообразование. В наших экспериментах при бензоналовой индукции наблюдалось статистически значимое увеличение содержания как свободного Т3, так и свободного Т4, что, вероятно, и является основой усиления биосинтетических процессов, приводящих к гипертрофии субклеточных структур гепатоцитов и усиления синтеза белка.
Известно, что синтез, секреция и действие гормонов щитовидной железы контролируется гипоталамо-гипофизарно-тиреоидной системой. Тиреотропин-рилизинг-фактор, секретируемый гипоталамусом, стимулирует синтез и секрецию тиреотропного гормона. В наших исследованиях именно при бензоналовой индукции наблюдалась тенденция к повышению уровня ТТГ. Зиксориновая индукция сопровождалась повышением уровня только свободного Т3. Содержание свободного Т4 даже имело тенденцию к снижению. При этом такую же тенденцию имело содержание ТТГ.
В исследовании для ингибирования монооксигеназной системы печени были использованы «эталонные» ингибиторы микросомального окисления – хлорид кобальта и циметидин. Хотя оба вещества и относятся к ингибиторам монооксигеназной системы печени, однако они совершенно различаются по химическому строению. Кобальт – необходимый элемент для нормального функционирования организма, так как он входит в состав ряда ферментов. Однако высокие дозы данного элемента токсичны [6]. Он проявляет генотоксичность [7], вызывает окислительный стресс [8] и гипоксию [9]. В то же время молекулярные механизмы токсичности кобальта до конца не ясны. Снижение содержания микросомальных цитохромов Р-450 и b5 при введении солей кобальта, возможно, связано с индукцией гемоксигеназы [10]. Возможна также инактивация цитохрома Р-450 в цитохром Р-420 при интоксикации солями кобальта за счёт фосфорилирования его белковой части [11].
Циметидин – N-циано-N’-метил-N»-гуанидин. Его эмпирическая формула – C10H16N6S. Это вещество является первым синтезированным блокатором гистаминовых Н2-рецепторов, за счёт чего ингибирует секрецию кислоты в желудке. Циметидин ингибирует НАДФН-зависимое микросомальное окисление, связанное с цитохромом Р-450 [12], что и приводит к уменьшению содержания восстановленного цитохрома Р-450 и значительному подавлению анилингидроксилазной активности монооксигеназ печени. Вероятно, именно различия в механизмах ингибирования и явились причиной разнонаправленного изменения содержания тиреоидных гормонов при хлоридкобальтовом и циметидиновом ингибировании. Для наглядности этих изменений проанализировали соотношения сТ3/Т3 и сТ4/Т4 при разных формах ингибирования монооксигеназной системы печени.
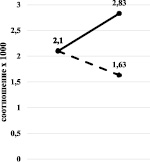
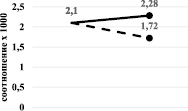
Результаты показали, что при ингибировании хлоридом кобальта наблюдается повышение соотношения как сТ3/Т3, так и сТ4/Т4 (рис. 4, а), а при циметидиновом ингибировании сохранение соотношения сТ3/Т3 (Р > 0,05) и снижение – сТ4/Т4 (рис. 4, б).
а) б)
Рис. 4. Изменение соотношения сТ3/Т3 (а) и сТ4/Т4 (б) при хлоридкобальтовом (сплошная линия) и циметидиновом (прерывистая линия) ингибировании
В обоих случаях изменения произошли на фоне снижения содержания ТТГ. Известно, что именно содержание ТТГ первым реагирует на нарушение функций щитовидной железы. Его понижение может проявляться на бессимптомных стадиях болезней, когда показатели Т3 и Т4 ещё в норме. В наших экспериментах на фоне повышения уровня Т3 и отчасти Т4 наблюдалось снижение уровня ТТГ и, в особенности, при ингибировании монооксигеназной системы хлоридом кобальта. Эти изменения, вероятно, указывают на нарушения функции щитовидной железы за счёт прямого (может быть в случае хлоридкобальтового ингибирования) или опосредованного (может быть в случае циметидинового ингибирования) воздействия на неё применённых ингибиторов.
Заключение
Таким образом, результаты исследований показали, что как индукция, так и ингибирование монооксигеназной системы приводит к повышению содержания тиреоидных гормонов. Однако при индукции наблюдается не изменение или тенденция к повышению уровня ТТГ, а при ингибировании его статистически значимое снижение. Полученные результаты свидетельствуют о наличии зависимости между функциональным состоянием монооксигеназной системы печени и содержанием тиреоидных гормонов в организме. Однако часть результатов можно объяснить с позиции индивидуального влияния использованных препаратов. Исходя из этого можно заключить, что зависимость между функциональным состоянием монооксигеназной системы и тиреоидным статусом является не прямой, а опосредованной.
Библиографическая ссылка
Юлдашев Н.М., Расулова М.Т. ТИРЕОИДНЫЙ СТАТУС ОРГАНИЗМА ПРИ ИНДУКЦИИ И ИНГИБИРОВАНИИ МОНООКСИГЕНАЗНОЙ СИСТЕМЫ ПЕЧЕНИ В ЭКСПЕРИМЕНТЕ // Научное обозрение. Биологические науки. 2019. № 4. С. 26-32;URL: https://science-biology.ru/en/article/view?id=1168 (дата обращения: 23.06.2026).
DOI: https://doi.org/10.17513/srbs.1168