

 science-review.ru
science-review.ru
Scientific journal
Scientific Review. Biological science
ISSN 2500-3399
ПИ №ФС77-57454
THE REGENERATIVE POTENTIAL OF THE ORGANISM WITH CONTROLLED MODERATE RESPIRATION

Поступающий с дыханием в организм кислород О2 определяет энергетические возможности биологических существ. Каждому виду млекопитающих присуща своя интенсивность дыхательного метаболизма: мыши потребляют до 900 мм3 кислорода О2 за час на 1 г веса, слон около сотни; для человека этот параметр составляет ~ 200 мм3 О2 ∙г-1∙ч-1 [1].
Пребывание в атмосфере с пониженной концентрацией кислорода стимулирует процессы адаптации к гипоксии, что при научно обоснованном методическом применении создает условия для оздоровления и активного долголетия [2].
Из газоразрядной аналогии для неравновесных систем полагается, что интенсивность кислородного обмена влияет на устойчивость организма к заболеваниям энергетической природы [3]. Тем самым контролируемое умеренное дыхание способно увеличить восстановительный потенциал организма.
Цель – исследовать влияние умеренного дефицита кислорода на состояние человеческого организма с точки зрения термодинамического подхода.
Особенности потребления энергоресурсов для различных биоорганизмов
Зависимости потребления кислорода О2 при дыхании возможно связать с некоторыми энергетическими закономерностями [4]. При этом интенсивность метаболизма представляется по энергонасыщенности V и потреблению энергоресурсов i в организме человека и различных по размерам животных как ряд зависимостей V(i) (рис. 1).
Для сравнения весьма различающихся биоорганизмов величины V и i могут быть рассмотрены для единичного объема, в условно подобных точках внутри тела и схожей биоткани, к примеру мышечной.
Энергонасыщенность V представляется как возможность совершения работы, или «активность». Тем самым V может определяться, например, как сумма тепловых эффектов реакций (химической индукции и каталитических ферментативных), характерных для рассматриваемой биоструктуры, в данном объеме за единицу времени.
Потребление ресурсов i рассматривается преимущественно в аспекте дыхания, и для выделенного объема пропорционально потоку кислорода О2 через него. Абсолютное потребление кислорода О2 различными млекопитающими растет при увеличении массы тела. В соответствии с зависимостью [1], за час кролик усваивает несколько литров кислорода, человек 10–20 л, слон 400–500 л, и поток O2 из глубины тела наружу через выбранный объем больше для массивного животного.
В рассматриваемой области энергия потока i частично расходуется, и произведение (V·i) можно считать пропорциональным плотности выделяющейся мощности W.
В обычном нормальном состоянии относительного спокойствия (линия N) организм млекопитающих пребывает преимущественное время. Умеренное и далее глубокое гипоксическое состояние наступает по мере снижения количества поступающего в организм кислорода О2 (линия G). Состояние воспаления и начала перерождения биотканей соответствует энерговыделению, превышающему критическое Wcr = Vcr · icr = const (линия С).
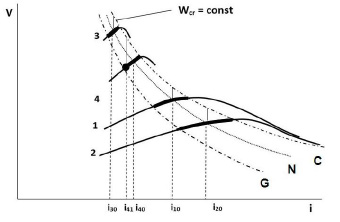
Рис. 1. Характеристики энергонасыщенности (активности) V для различных биоорганизмов в зависимости от потребляемых энергоресурсов i. N – нормальное состояние различных млекопитающих, G – гипоксическое состояние, C – граница критического перерождения биоструктур. 1 – человек, 2 – крупное млекопитающее, 3 – мелкое млекопитающее, 4 – млекопитающее Heterocephalus glaber («голый землекоп»)
Коридор жизненных состояний по величине удельного потребления кислорода i закономерно увеличивается от мелких к крупным млекопитающим (область между G и C).
Устойчивость биоструктур различных млекопитающих
Для каждого вида можно говорить о запасе устойчивости ξ биоструктур по отношению к их перерождению [4]. При появлении дефекта и торможения на нем энергопотока величина V может нарастать, приводя к образованию энергетической ступеньки – «градиента». Физиологически градиент проявляется в виде области из воспаленной биоткани, в которой повышается энерговыделение за счет, например, термогенеза.
Оценочно для конкретного уровня потребления кислорода i значение ξ соответствует разнице в энергонасыщенности V, определяющей переход от состояния на зависимости V(i) до линии C критического энерговыделения Wcr. С эволюционной точки зрения, ввиду сходной белковой структуры биоорганизмов, в обычном N состоянии величина ξ должна быть близкой по величине у различных млекопитающих, например для человека при i10, для крупного млекопитающего при i20 и мелкого при i30 (рис. 1, вертикальные сплошные линии).
У мелких млекопитающих интенсивность метаболизма в организме выше, что при появлении проблемной области провоцирует быстрое формирование энергоградиента. К тому же мелкие животные, как правило, чаще подвергаются стрессовым обстоятельствам, например угрозе жизни. Оба фактора соответствуют смещению вдоль энергетической зависимости V(i) от N к линии C, и при нахождении в данном состоянии запас устойчивости ξ снижается.
Тем самым чисто термодинамические условия функционирования организма млекопитающего накладываются на статистику его поведенческих особенностей. Например, процент онкологической патологии в популяции оказывается меньшим для крупных животных, таких как слоны или киты [5].
Млекопитающие «голые землекопы», обитающие в норах под землей, практически постоянно находятся в условиях пониженной концентрации кислорода [6]. Поэтому на зависимости 4 (рис. 1) выделенная точка обычного состояния Heterocephalus glaber располагается левее кривой N, характеризующей прочих млекопитающих, и соответствует потреблению кислорода i41 вместо условно нормального i40. Это обычное для «землекопов» умеренное гипоксическое состояние имеет повышенный запас устойчивости ξ, характеризуется возможностью репарации и регенерации биоструктур, резистентностью к онкологическим заболеваниям [7, 8]. В то же время у «землекопов», извлеченных в неестественные условия зоопарка или лаборатории, опухолеобразование становится возможным [9].
Постоянное легкое кислородное голодание в перспективе способствует долголетию: «голые землекопы» живут до 30 лет и более, что на порядок превышает продолжительность жизни равновеликих им млекопитающих.
Энергетические зависимости для живых организмов, отличающихся строением от млекопитающих, должны быть соответственно смещены. Например, ввиду менее интенсивного удельного метаболизма для хладнокровных животных гипербола Wcr должна располагаться ближе к началу координат (V, i).
Теории активного долголетия
В настоящее время широко известны гипотезы, указывающие в качестве причин старения изменения в генетическом аппарате, накопление мутаций, бактериальную интоксикацию, свободные радикалы [10]. Так, в подходе академика В.П. Скулачева старость представляется наступлением состояния, когда в организме число отмирающих клеток начинает превышать количество рождающихся, чему можно противодействовать повышением устойчивости клеточных структур, в особенности митохондрий [11, 12].
Влияние умеренного гипоксического воздействия на физиологические процессы в организме
Ввиду большей крутизны зависимости V(i) при малых i, изменению Δi соответствует значительный отклик по энергонасыщенности ΔV. Тем самым небольшие управляющие воздействия способны произвести значительную реконструкцию в биотканях.
Разнообразие возможных энергетических процессов обеспечивает адекватную реакцию организма к отклонению гомеостаза и внешним воздействиям, что означает повышенную эффективность иммунной системы защиты именно в состоянии гипоксического расслабления. Характерными являются активизация гипоксией индуцируемого фактора (HIF) и его влияние на иммунитет [13].
В физиологическом плане при контролируемом умеренном потреблении О2 происходит активизация внутриклеточного (тканевого) дыхания.
Термодинамика жизненных процессов в аспекте долголетия
Пределы роста и изменчивости каждого млекопитающего ограничены особенностями его структурной организации. Ввиду неуклонно провоцируемых внешней средой дефектов в организме, для поддержания жизни синтез ключевых биоструктур должен происходить опережающим темпом. В организме непрерывно осуществляется превращение получаемой из пищи энергии U в свободную энергию G и в связанную Q, так что U = G + Q. Свободная энергия G после производства структурных преобразований в организме и совершения работы переходит в связанную форму Q и в итоге выделяется в виде тепла.
Можно принять, что для высокоорганизованного человеческого организма жизнь поддерживается, пока происходит производство энергии Gi, представляющей собой ту часть свободной энергии G, которая обеспечивает протекание реакций химической индукции для синтеза молекул АТФ, нуклеиновых кислот ДНК и других сложных биоструктур.
Степень неравновесности, взятая как отношение (Gi / U), максимальна при рождении, поскольку в этот период имеет место активный рост и развитие органов (рис. 2). По прошествии времени, когда основные структуры организма сформированы и развиты, энергия требуется преимущественно для стабильного поддержания в них физиологических процессов. Так, интенсивность энергопотока в спокойном состоянии у новорожденных составляет 300 кДж·м-2·ч-1, в то время как у людей зрелого возраста – 120 кДж·м-2·ч-1 [14]. С возрастом доля созидающих процессов синтеза уменьшается, преобладающими остаются процессы потребления с участием ферментативных реакций. Соответственно, уменьшается число реакций химической индукции и снижается степень неравновесности. Когда величина (Gi / U) опускается ниже критической величины (Gi / U)cr, определенной для каждого организма, жизнь прекращается.
При выполнении гипоксической процедуры, в состоянии кислородного дефицита и физической расслабленности, внутренняя энергия U в меньшей степени тратится на совершение физической работы и прочие процессы, определяющие связанную энергию Q. Степень неравновесности (Gi / U) несколько увеличивается, и ее влияние можно считать пролонгированным на продолжительный срок (кривая 1). Соответственно увеличивается доля свободной энергии G, направляемой на процессы синтеза, в том числе для целей регенерации.
В случае заболевания, имеющего энергетическую причину, в организме развиваются процессы, сдвигающие химическое равновесие к угнетению реакций индукции, и срок жизни сокращается (кривая 2).
Влияние гипоксических процедур на продолжительность жизни
Процедуры, позволяющие достичь умеренной гипоксии и физической релаксации в организме, обладают значительным восстановительным потенциалом.
С течением времени в тканях организма неизбежно накапливаются дефекты с интенсивностью N не менее определенной, например, ввиду наличия естественного радиоактивного фона. При накоплении некоторой критической плотности дефектов N* наступает деградация и гибель организма, ввиду затрудненного энергопотока и снижения производства свободной энергии. Закон накопления дефектов имеет характерный экспоненциальный вид N ~ N’·exp(A·t), где N’ – уровень дефектов при рождении (рис. 3). Временной коэффициент соответствует величине А = ln (N*/N’) / T, где Т – средняя продолжительность жизни, обусловленная наследственностью, условиями жизни, питания и т.д. Начальный уровень N’ для любых биоорганизмов можно считать относительно невысоким по сравнению с критическим N*.

Рис. 2. Изменение степени неравновесности (Gi /U) человека в течение времени t жизни. 1 – случай выполнения одиночной гипоксической процедуры, 2 – проявление заболевания энергетической природы. (Gi /U)cr – критический для жизни уровень неравновесности
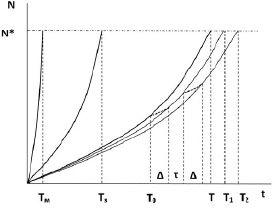
Рис. 3. Накопление числа дефектов N в организме в течение времени t жизни. N* – критическая плотность дефектов в организме.
Продолжительность жизни: Тм – мышь, Тз – «голый землекоп», Т – человек. Т0 – начало гипоксического тренинга, Т1 – сдвиг линии жизни человека после выполнения одиночной гипоксической процедуры. ∆ – время действия положительного эффекта от гипоксической процедуры, τ – промежуток между процедурами
Чтобы накопление деструктивных элементов не препятствовало протеканию энергопотоков в организме, требуется организовать по крайней мере периодические гипоксические восстановительные процедуры, которые воссоздают функциональность биотканей с нарушенной структурой.
Для мелкого млекопитающего типа мыши продолжительность жизни составляет Тм ~ 2–3 года (рис. 3). В то же время схожий по размерам «голый землекоп» живет до Тз ~ 30 лет. Если в данном случае эффект долгожительства отнести на счет умеренного кислородного голодания и связанной с этим реабилитационной активностью, то и для человеческого организма как подобной белковой структуры можно ожидать роста продолжительности жизни Т при периодическом воздействии контролируемой гипоксии.
В условиях санатория или медицинского стационара возможность погружения в умеренную гипоксию доступна на оборудовании типа гипоксических камер или кабин. В повседневных условиях тренинг возможен в виде периодических дыхательных процедур, в частности «произвольного зевания», при которой путем продолжительной задержки дыхания после выдоха достигается умеренная гипоксия при эффективном расслаблении организма, а за счет замены вдоха зевком обеспечивается контроль оптимального режима [3].
Восстановительный эффект гипоксической процедуры
Можно представить, что умеренное гипоксическое воздействие восстанавливает часть дефектных биоструктур и уменьшает скорость накопления дефектов в организме, поскольку разрывает зависимость порождения одними дефектами других. При этом каждая выполненная гипоксическая процедура схематично переводит организм на «линию жизни» с большей продолжительностью.
Для наглядности удобно перестроить рис. 3 к полулогарифмическим координатам «ln N – t», при этом экспоненциальные зависимости заменятся на прямые линии. Участок экспоненты за период ∆ можно для оценки заменить горизонтальным отрезком, как если бы число дефектов оказалось зафиксированным в ходе процедуры. При этом прирост продолжительности жизни от одиночной гипоксической тренировки, рассматриваемый как переход организма из исходного состояния («линия жизни» Т) в состояние с оздоровительным эффектом (линия Т1), можно оценить для этой первичной процедуры как (Т1 – Т) = Т / Т0 · ∆. Переход по линиям жизненного цикла Т…Т1…Т2… для оценки можно принять пропорциональным количеству процедур n = (Т–Т0) / (∆+τ). В итоге к статистически ожидаемому сроку жизни Т прирост продолжительности за счет периодического выполнения дыхательных занятий составит
(Тn –Т) ~ (Т1 – Т) · n = Т·(Т/Т0 –1)/(1+ τ/∆).
К примеру, примем, что наследственная предрасположенность человека составляет Т ~ 70 лет, гипоксические тренировки начались с возраста Т0 = 20 лет. Время продолженного влияния от гипоксической процедуры можно оценить величиной по крайней мере ∆ = 1 ч. Если периодичность тренинга составляет 1 раз в сутки, то τ ~ 23 ч. При этих условиях можно ожидать прибавки продолжительности жизни (Тn –Т) ~ 7 лет, или 10 %.
Приведенная оценка в значительной мере условна, но демонстрирует основные тенденции повышения восстановительной способности организма через умеренные гипоксические процедуры.
При регулярном выполнении гипоксических процедур можно рассчитывать на эффект «тренированности» в виде снижения интенсивности накопления дефектов в организме.
Устойчивость биотканей к перерождению у млекопитающих зависит от метаболизма, определяемого интенсивностью дыхания. При этом на характер газообмена у различных видов млекопитающих существенно влияют их поведенческие особенности.
Умеренное гипоксическое состояние для человеческого организма способствует повышению устойчивости биоструктур. Условие гипоксического расслабления соответствует разнообразию возможных энергетических процессов в организме, что обеспечивает эффективную регенерацию тканей и активное долголетие.
В целях профилактики можно рассчитывать на периодическую гипоксическую дыхательную процедуру, в частности контролируемой «произвольной зевоты», обеспечивающую режим оптимального перехода в состояние восстановительной релаксации.
Оценочно в случае регулярного гипоксического тренинга для статистически предполагаемого срока жизни 70–80 лет можно ожидать увеличения периода активного долголетия на 10 % и более.
Библиографическая ссылка
Шушков С.В. ВОССТАНОВИТЕЛЬНЫЙ ПОТЕНЦИАЛ ОРГАНИЗМА ПРИ КОНТРОЛИРУЕМОМ УМЕРЕННОМ ДЫХАНИИ // Научное обозрение. Биологические науки. 2022. № 1. С. 62-67;URL: https://science-biology.ru/en/article/view?id=1262 (дата обращения: 28.07.2026).
DOI: https://doi.org/10.17513/srbs.1262