

 science-review.ru
science-review.ru
Scientific journal
Scientific Review. Biological science
ISSN 2500-3399
ПИ №ФС77-57454
SPATIAL AND AGE STRUCTURE OF CENOPOPULATIONS OF CORYDALIS SOLIDA (L.) CLAIRV

В качестве объекта исследования выбран весенний эфемероид – Coridalis solida (L.) Clairv – Хохлатка плотная (семейство Fumariaceae). Данный вид занесен в Красную книгу Архангельской области и находится в категории 3 – редкий вид. В условиях области группа растений, относящихся к эфемероидам, относительно слабо изучена, в первую очередь из-за кратковременного вегетационного периода и труднодоступности территории в весенний период из-за стаивания снежного покрова. В силу упомянутых выше причин изучение эфемероидов является актуальным. На территории Беломорского заказника выявлено несколько точек произрастания эфемероидов, где провели исследования по изучению пространственной и возрастной структуры ценопопуляций, в частности в окрестностях с. Вознесенье, где и были заложены трансекты и проводился учет.
Беломорский государственный природный биологический заказник регионального значения был создан в 1998 году и до сих пор выполняет свои функции ООПТ: изучение редкой флоры в пределах особо охраняемых природных территорий.
Вид Corydalis solida – это растение подсемейства Дымянковые, которое относится к эфемероидам. Эфемероиды – многолетние травянистые полициклические растения с коротким циклом жизни надземных побегов, приходящимся на весну, и длительным периодом относительного покоя в летний сезон. В эту экологическую группу входят представители разных жизненных форм и таксономических групп [1].
Хохлатка растет на опушках лиственных лесов, в ольшаниках, зарослях кустарников, на лужайках, пойменных и суходольных лугах. Предпочитает рыхлые, богатые гумусом почвы. По отношению к влаге хохлатка относится к мезофитам, по отношению к освещенности – факультативный гелиофит, по отношению к трофности почвы – мезотроф. Активная фаза цикла приурочена к апрелю-маю [2].
При изучении теоретического материала по онтогенезу Corydalis solida были выявлены диагностические признаки возрастных состояний – строение листовой пластинки и наличие соцветий. В ходе полевых исследований проведена работа по выявлению возрастных состояний хохлатки в природных ценопопуляциях, в частости на территории ООПТ «Беломорский заказник». Территория исследования – это пойменные луга и закустаренные участки, расположенные по краю сельского кладбища. Почвы везде богатые и влажные, что и характерно для данного вида.
Цель работы: изучить пространственную и возрастную структуру ценопопуляции Corydalis solida в пределах фитоценозов с разной степенью освещенности.
Материалы и методы исследования
Для определения возрастной и пространственной структур были заложены 2 трансекты [3]: в затенении, в пределах закустаренного сообщества по краю кладбища и на открытом пространстве луга. Каждая трансекта разбивалась на 15 учетных площадок (50×50 см). В пределах каждой учетной площадки подсчитывались особи всех возрастных состояний. У генеративных особей определялось количество цветов в соцветиях. На основе полученных данных определялись пространственная структура и возрастная структура ценопопуляций [4].
Возрастные состояния данного вида были изучены по онтогенезу О.В. Смирновой и В.А. Черемушкной (1975 г.). Выявлены диагностические отличия возрастных состояний:
1) проростки (p) – имеют единственную семядолю и длинный черешок;
2) ювенильные (j) – имеют 1 срединный ассимилирующий лист и чаще всего 2 чешуевидных листа;
3) имматурные (im) – одна листовая пластинка трижды тройчато-рассеченная;
4) виргинильные (v) – два срединных листа, две листовые пластинки трижды тройчато-рассеченные;
5) молодые генеративные (g1) – имеют один генеративный побег;
6) средневозрастные генеративные (g2) – 2–4 генеративных побега. Дополнительный генеративный побег развивается из почки в пазухе верхнего чешуевидного листа;
7) старые генеративные (g3) – особи-раметы, 1 генеративный побег;
8) сенильные (s) – верхушечная или заменившая ее пазушная почка формируют 1–2 чешуевидных и 1 срединный лист однажды тройчато-рассеченный, ювенильного типа.
У экземпляров с нормальной или пониженной жизненностью онтогенез заканчивается в фазе одноосного побега, у экземпляров с повышенной жизненностью формируется недолговечный клон, состоящий обычно из старых вегетативных и сенильных особей (рис. 1) [5].
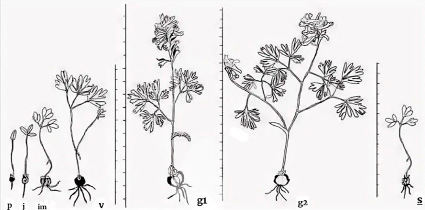
Рис. 1. Возрастные состояния Corydalis solida [6]
Таблица 1
Численность ценопопуляции Corydalis solida в затенении
|
№ площадки |
Возрастное состояние |
Общая численность, шт. |
|||||||
|
p |
j |
im |
v |
g1 |
g2 |
g3 |
s |
||
|
1 |
40 |
57 |
36 |
29 |
16 |
9 |
0 |
0 |
187 |
|
2 |
76 |
51 |
36 |
25 |
8 |
4 |
1 |
0 |
201 |
|
3 |
49 |
31 |
27 |
11 |
11 |
4 |
0 |
0 |
133 |
|
4 |
92 |
24 |
25 |
5 |
3 |
7 |
1 |
0 |
157 |
|
5 |
79 |
31 |
28 |
12 |
5 |
13 |
3 |
0 |
171 |
|
6 |
35 |
15 |
19 |
9 |
2 |
12 |
0 |
0 |
92 |
|
7 |
12 |
14 |
8 |
15 |
6 |
11 |
1 |
1 |
68 |
|
8 |
52 |
12 |
9 |
10 |
6 |
13 |
0 |
0 |
102 |
|
9 |
53 |
14 |
10 |
10 |
4 |
10 |
3 |
1 |
105 |
|
10 |
66 |
27 |
5 |
4 |
2 |
13 |
2 |
0 |
119 |
|
11 |
52 |
20 |
7 |
10 |
1 |
12 |
1 |
0 |
103 |
|
12 |
89 |
5 |
1 |
6 |
4 |
9 |
2 |
0 |
116 |
|
13 |
22 |
13 |
25 |
8 |
2 |
4 |
1 |
2 |
77 |
|
14 |
12 |
4 |
5 |
3 |
2 |
8 |
0 |
0 |
34 |
|
15 |
21 |
7 |
8 |
6 |
3 |
5 |
1 |
0 |
51 |
|
Всего особей |
750 |
325 |
249 |
163 |
75 |
134 |
16 |
4 |
1716 |
|
% |
42,6 |
18,9 |
14,5 |
9,5 |
4,4 |
7,8 |
0,9 |
0,2 |
100% |
|
Ср. число особей на площадке |
50 |
21,7 |
10,9 |
8,9 |
16,6 |
5 |
1,1 |
0,3 |
114,4 |
|
Число особей на 1 м2 |
50 |
21,7 |
10,9 |
8,9 |
16,6 |
5 |
1,1 |
0,3 |
|
Результаты исследования и их обсуждение
При камеральной обработке рассчитывали численность и плотность ценопопуляции хохлатки, а также численность особей всех ее состояний. Средние данные отображены в таблице 1.
Всего на целой трансекте получилось 1716 особей разных возрастных состояний. По количеству преобладают проростки – 750 особей. Меньше всего сенильных – 4. Данные значения по возрастным состояниям наблюдаются в общем на всех площадках – количество особей равномерно распределено по территории. В пределах площадок больше всего особей на 2-й учетной площадке – 201, меньше всего на 14-й – 34 особи. Подобный разброс, скорее всего, связан с тем, что трансекту закладывали случайным образом и часть ее попала на чрезмерно закустаренную область, следовательно, подходящих мест для произрастания хохлатки было меньше.
Коэффициент вариации более 100% указывает на наличие значений, сильно отличающихся от средней величины. В исследуемой совокупности сильна вариация признаков по отношению к средней величине. Получены следующие показатели: 53% (p), 72% (j), 71% (im), 67% (v), 81% (g1), 38% (g2), 96% (g3), 222% (s). Рассеивание данных относительно точечной оценки математического ожидания высокое у g2, p. Очень высокий разброс относительно выборочного среднего у v, im, j, g1, g3. Аномальный разброс s обусловливается маленьким объемом выборки. Все выборки неоднородны, что вызвано недостаточным объемом выборки и образованием статистических выбросов площадками 7, 13, 14, 15. Наиболее высокой коэффициент вариации по численности особей между площадками равен 42%.
На основании таблицы 1 построим диаграмму возрастного спектра ценопопуляции (рис. 2).
Базовый спектр ценопопуляции в затенении демонстрирует левосторонний спектр с преобладанием особей на ранней стадии онтогенеза (рис. 2). Данный спектр является характерным для хохлатки, об этом свидетельствует информация из литературных источников (Смирнова, 1987). Такой спектр принято называть инвазионным, т.е. находящимся на стадии внедрения в данный фитоценоз. Возможно, этому способствуют экологические особенности вида, так как хохлатка является факультативным гелиофитом, мезофитом и эвтрофом. 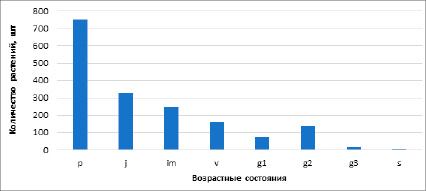
Рис. 2. Возрастной спектр ценопопуляции Corydalis solida в затенении
Таблица 2
Численность ценопопуляции Corydalis solida на открытом пространстве
|
№ площадки |
Возрастное состояние |
Общая численность, шт. |
|||||||
|
p |
j |
im |
V |
g1 |
g2 |
g3 |
s |
||
|
1 |
2 |
3 |
17 |
11 |
1 |
0 |
0 |
0 |
34 |
|
2 |
11 |
5 |
7 |
2 |
3 |
1 |
0 |
0 |
29 |
|
3 |
3 |
1 |
5 |
3 |
2 |
0 |
0 |
0 |
14 |
|
4 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
|
5 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
|
6 |
2 |
1 |
0 |
0 |
0 |
0 |
0 |
0 |
3 |
|
7 |
4 |
3 |
5 |
0 |
0 |
0 |
0 |
0 |
12 |
|
8 |
0 |
2 |
0 |
0 |
0 |
0 |
0 |
0 |
2 |
|
9 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
|
10 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
|
11 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
|
12 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
|
13 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
|
14 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
|
15 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
|
Всего особей |
22 |
15 |
34 |
16 |
6 |
1 |
0 |
0 |
94 |
|
% |
23,4 |
15,96 |
36,17 |
17,03 |
6,38 |
1,06 |
0 |
0 |
100% |
|
Ср. число особей на площадке |
1,5 |
1 |
2,27 |
1,07 |
0,4 |
0,07 |
0 |
0 |
|
|
Число особей на 1 м2 |
1,5 |
1 |
2,27 |
1,07 |
0,4 |
0,07 |
0 |
0 |
|
Далее рассчитываем численность и плотность популяции на открытом пространстве и численность особей всех состояний. Данные представлены в таблице 2.
Всего на данной трансекте было 94 особи C. solida. Больше всего проростков и особей имматурного возрастного состояния. Старогенеративных и сенильных возрастных состояний не было. Максимальная численность растений – 34, минимальная – 2. Коэффициент вариации возрастных состояний следующий: 201% (p); 156% (j); 208% (im); 271% (v); 228% (g1); 387% (g2); коэффициент вариации по общей численности 179%. Обнаружен аномальный разброс данных относительно выборочного среднего. Высокие показатели коэффициента вариации связаны с очень неравномерным распространением особей по учетным площадкам, нерепрезентативным объемом выборки, статистическими выбросами. На большинстве площадок особей не наблюдалось. Возможно, это связано с тем, что на лугу выражен явный микрорельеф: есть переувлажненные понижения и повышения. Также высока плотность злаков, которая способствует образованию плотной дернины на лугу, мешающей распространению данного вида.
На основании таблицы построим диаграмму возрастного спектра ценопопуляции (рис. 3)
Перед нами точно такой же тип спектра, что и для первой ценопопуляции. Для всех видов хохлаток характерны высокая экологическая пластичность по отношению к факторам освещенности и способность внедряться в различные виды местообитаний [7]. Но, так как хохлатка – это эвтроф и ей нужны богатые почвы, а также высокая влажность, то два этих фактора могут компенсировать ее рост на открытом пространстве [8].
На последнем этапе определяли тип пространственного размещения особей на двух трансектах. Для этого по методике проводили математические вычисления. Результаты и средние показатели представлены в таблице 3.
Так как в обоих случаях σ² > m, распределение получилось групповое, что также доказывает коэффициент вариации. Литературные источники также подтверждают, что для хохлатки характерно групповое распределение [9].
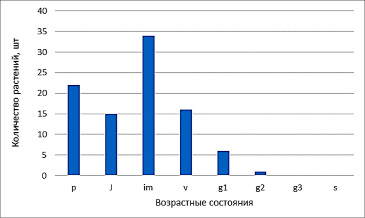
Рис. 3. Возрастной спектр ценопопуляции Corydalis solida на открытом пространстве
Таблица 3
Тип пространственного размещения Corydalis solida
|
Средние показатели |
|
|
В затенении |
На открытом пространстве |
|
σ2 = Σ(m – V)2 / (N – 1), где m – среднее число особей на площадке. V – число особей на конкретной площадке. N – число площадок |
|
|
M = 113,9 |
M = 94 |
|
Σ(m – V)2 = 33048,95 |
Σ(m – V)2 =37694 |
|
σ2 = 2360,64 |
σ2 = 2692,43 |
Заключение
В ходе исследования изучены пространственная и возрастная структуры ценопопуляций Corydalis solida. Популяция Corydalis solida в затенении является устойчивой, что связано с особенностями местопроизрастания: более оптимальные богатые почвы, дренируемые кустарниками и деревьями. Популяция, изученная на открытом пространстве, не устойчива. Скорее всего, это связано с выраженным микрорельефом на лугу, закочкаренностью, а также с высокой влажностью. Распределение в затенении и на открытом пространстве групповое. Всхожесть на обеих трансектах высокая, но при этом это косвенно указывает на низкую выживаемость проростков при переходе на следующее возрастное состояние.
Библиографическая ссылка
Тюкачёва А.А., Амосова И.Б. ПРОСТРАНСТВЕННАЯ И ВОЗРАСТНАЯ СТРУКТУРА ЦЕНОПОПУЛЯЦИИ CORYDALIS SOLIDA (L.) CLAIRV // Научное обозрение. Биологические науки. 2023. № 3. С. 11-16;URL: https://science-biology.ru/en/article/view?id=1330 (дата обращения: 03.07.2026).
DOI: https://doi.org/10.17513/srbs.1330