

 science-review.ru
science-review.ru
Scientific journal
Scientific Review. Biological science
ISSN 2500-3399
ПИ №ФС77-57454
DIET FEATURES OF THE EURASIAN EAGLE OWL (BUBO BUBO) IN TRANSFORMED ARALQUM DESERT ECOSYSTEMS

Введение
Аралкум представляет собой молодую пустыню, сформировавшуюся во второй половине XX в. на месте высохшего дна Аральского моря вследствие резкого понижения уровня воды. Данный регион расположен преимущественно в пределах Республики Каракалпакстан (Узбекистан), а также в южной части Казахстана, формируя в природно-географическом отношении новый ландшафтный комплекс Приаралья.
Рельеф Аралкума преимущественно равнинный и слабоволнистый, сложенный аккумулятивными морскими отложениями бывшего дна. Широко распространены солончаки и песчаные массивы; в ряде участков под воздействием эоловых процессов сформировались барханы и песчаные гряды. Абсолютные высоты колеблются в пределах 30–60 м над уровнем моря. В геоморфологическом отношении территория остается нестабилизированной: активно протекают процессы дефляции, сопровождающиеся переносом солевых и пылевых частиц на значительные расстояния [1].
Растительный покров относится к пустынному типу и представлен преимущественно галофитными и псаммофитными видами. На ранних этапах формирования пустыни растительность отличалась крайней разреженностью, однако по мере развития сукцессионных процессов наблюдается расширение ареалов ксеро- и галофитных сообществ. Основу фитоценозов составляют саксаул (в особенности черный саксаул), кандим (Calligonum), янтак (Alhagi), солянки (Salsola) и тамарикс (Tamarix) [2].
Аралкум является одним из наиболее молодых и в то же время наименее изученных природных регионов Узбекистана. Его площадь продолжает увеличиваться по мере дальнейшей регрессии Аральского моря, что обусловливает динамичный характер формирования фауны и трансформацию экосистемных связей. Наибольшее видовое разнообразие приурочено к территориям бывших островов – Возрождения, Комсомольский, Константина, Лазарева и Беллинсгаузена, а также к ряду менее крупных островных участков, которые после обмеления моря соединились с материком. Систематическое изучение фауны этих изолированных в прошлом территорий началось в начале 2010-х гг., после их фактического соединения с сушей и снятия ограничений на посещение [3].
Современные исследования показывают, что экосистемы Аралкума находятся в стадии активного формирования и характеризуются высокой динамичностью природных процессов. По данным I. S. Plotnikov, N. V. Aladin (2023), на осушенном дне Аральского моря постепенно формируется новый пустынный природный комплекс с измененной структурой флоры и фауны [4]. Исследование J. P. Deroin (2025) подтверждает продолжающиеся процессы опустынивания и трансформации ландшафтов региона [5]. В отчете United Nations Development Programme также отмечается постепенная адаптация животных и растений к новым экологическим условиям Аралкума [6].
По современным данным, на территории Аралкума зарегистрировано более 160 видов птиц, адаптирующихся к новым условиям трансформированной пустынной экосистемы [7; 8]. В частности, в исследованиях, проведенных на территории Казахстан, а также в ряде фаунистических работ, посвященных изучению животного мира региона, зарегистрировано 165 видов птиц [9; 10]. Из отряда совообразных здесь обитают филин Bubo bubo и домовый сыч Athene noctua. Оба вида являются обычными оседлыми представителями местной авифауны.
В работе Е. А. Быковой и соавт. (2020) впервые был проведен сравнительный анализ питания хищных птиц (Bubo bubo, Athene noctua и Aquila chrysaetos) в различных биотопах бывшего острова Возрождения и сопредельных районов; однако основное внимание уделялось позвоночным животным, вследствие чего полный спектр кормовых объектов не был рассмотрен [11]. В работах Ж. М. Ёркулова и др. (2024) относительно широко освещен рацион питания этой хищной птицы [12].
Исследование рациона филина как оседлого хищника Аралкума имеет важное научное значение для понимания формирования трофических связей в условиях молодой пустынной экосистемы и оценки направлений ее дальнейшей сукцессии.
Цель исследования – изучение трофической биологии филина в условиях Аралкума, поскольку данный аспект остается недостаточно разработанным для рассматриваемой территории.
Материалы и методы исследования
Полевые исследования проводились на территории пустыни Аралкум – молодой природной системы, сформировавшейся под влиянием антропогенной трансформации акватории Аральского моря. Район характеризуется высоким разнообразием мелких млекопитающих (прежде всего грызунов), рептилий, а также наземных и околоводных птиц, что формирует благоприятную кормовую базу для крупного хищника – филина.
В качестве основного материала исследования использованы погадки филина. Сбор проводился в 2024–2025 гг. в пяти постоянных местах гнездования вида на коренных островах и осушенном дне бывшего Аральского моря в поздневесенний (май) и осенний (ноябрь) сезоны (рис. 1). Координаты и особенности мест гнездования представлены в табл. 1. Все обследованные стации были условно разделены на три типа по биотопическому признаку: обрывы чинков и равнинные биотопы (солончаковая и закрепленная песчаная пустыня).
Цель исследования – оценка общего состава рациона филина, без анализа сезонной или межгодовой динамики. Полевой материал был собран в ходе двух экспедиционных выездов, после чего объединен в единую выборку для последующего анализа.
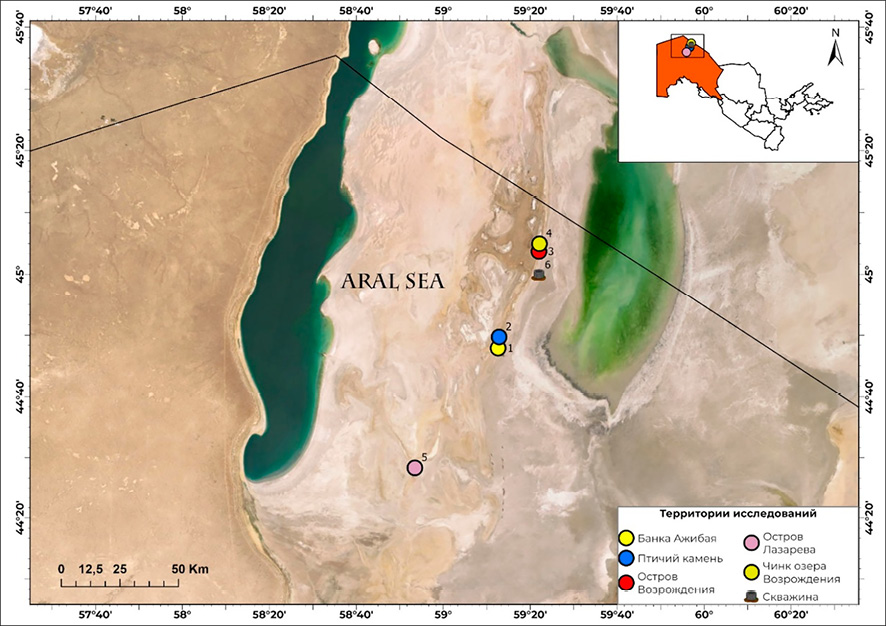
Рис. 1. Места сбора материала – гнезда филина Примечание: составлен авторами по результатам данного исследования
Таблица 1
Географические координаты и характеристика гнездовых участков филина
|
Участок |
Координаты |
Биотоп |
Краткая характеристика |
|
1. Банка Ажибая |
44.7981, 59.2106 |
Солончаковая пустыня |
Осушенное дно Аральского моря с каменистыми выходами |
|
2. Птичий камень |
44.82824, 59.21437 |
Солончаковая пустыня |
Каменистые участки с трещинами и пустотами |
|
3. Остров Возрождения, бухта Комсомольская |
45.0623, 59.3664 |
Обрывы чинка |
Абразионный береговой уступ (чинк) бывшего острова |
|
4. Остров Возрождения, озеро Возрождения |
45.08255, 59.36871 |
Обрывы чинка |
Чинковый участок рядом с сезонным озером |
|
5. Остров Лазарева |
44.472728, 58.891553 |
Закрепленные пески |
Песчаные массивы с выходами песчаника |
Примечание: составлена авторами на основе полученных данных в ходе исследования.
Таблица 2
Пространственное распределение погадок и жертв филина по участкам
|
Участок |
Дата сбора |
Количество погадок |
Количество жертв, особи |
|
1. Банка Ажибая |
06.05.2024 |
5 целых и 3 фрагмента |
12 |
|
2. Птичий камень |
07.05.2024 |
18 целых, 10 фрагментов и 2 разрозненных костных остатка |
66 |
|
3. Остров Возрождения, бухта Комсомольская |
07.05.2024 |
29 целых, 13 фрагментов и 4 разрозненных костных остатков |
83 |
|
4. Остров Возрождения, озеро Возрождения |
09.05.2024 |
46 целых, 20 фрагментов и 11 разрозненных косных остатков |
187 |
|
5. Остров Лазарева |
02.11.2025 |
10 целых, 4 фрагмента |
34 |
Примечание: составлена авторами на основе полученных данных в ходе исследования.
Каждая проба маркировалась индивидуально с указанием даты, географических координат и типа биотопа, после чего помещалась в бумажные пакеты и транспортировалась в лабораторию. Фрагментированные погадки и разрозненные костные остатки учитывались отдельно с четким указанием даты и места сбора в целях избегания двойного подсчета для корректной оценки количества погадок и содержащихся в них остатков добычи. Всего было собрано и обработано 108 целых и более 50 фрагментированных погадок, а также 17 остатков добычи из полностью разрушенных погадок. Объем материала и количество идентифицированных жертв представлены в табл. 2.
В лабораторных условиях погадки предварительно высушивались при естественной температуре, затем подвергались механической разборке с использованием игл и пинцетов. Костные остатки промывались, повторно высушивались и сортировались по морфологическим признакам.
Идентификация остеологического материала проводилась с применением определителей [13–15] и сопоставления с эталонными образцами, хранящимися в фондах зоологической коллекции Института зоологии АН РУз (Ташкент, Узбекистан).
Таксономическая принадлежность устанавливалась по возможности до вида или рода. В качестве диагностических элементов преимущественно использовались верхние (maxilla) и нижние челюсти (mandibula) позвоночных животных, а также части посткраниального скелета – плечевые (os humerus), бедренные (os femoris) кости и другие парные элементы скелета. Определение беспозвоночных животных проводилось специалистами-энтомологами Института зоологии АН РУз по сохранившимся остаткам хитинового покрова (кутикулы) и также хелицер, хитинизированных отростков. Ряд беспозвоночных был определен до рода из-за выраженной фрагментации останков, недостаточной для точной видовой идентификации.
Количественная оценка добычи проводилась по методу минимального числа особей (MNI – Minimum Number of Individuals). Для каждого таксона определяли наиболее часто встречающийся парный диагностический элемент скелета (например, правую или левую mandibula), при этом максимальное число однотипных элементов принималось за минимальное число особей данного вида:
MNI = max(R,L),
где R – число костей правой стороны;
L – число костей левой стороны.
Результаты исследования и их обсуждение
В изученных погадках были обнаружены останки 382 животных, относящихся к 34 видам (родам) позвоночных и беспозвоночных животных, видовой состав и количественные показатели которых представлены в табл. 3.
Поскольку специфика рациона питания хищных птиц связана с особенностями биoтопических условий той территории, в которой они обитают, был проведен биотопический анализ приуроченности видов жертв. Как указывалось выше, авторы условно разделили все обследованные стации на три основных биотопа: обрывы чинков, солончаковую и песчаную пустыни. Анализ биoтопической приуроченности видов жертв показал, что более высокое видовое богатство характерно для чинков и прилежащих территорий. Здесь было обнаружено 11 видов млекопитающих, 9 видов птиц, 2 вида рептилий и 10 видов беспозвоночных, всего 32 вида.
Таблица 3
Состав пищевого рациона филина по результатам анализа погадок на территории Аралкума
|
Виды жертв |
Место сбора материала |
Всего |
||||||||||||
|
1 |
2 |
3 |
4 |
5 |
Кол-во, особей (N) |
% от общего кол-ва особей |
||||||||
|
Беспозвоночные |
||||||||||||||
|
Каспийский галеод Galeodes caspius |
6 |
6 |
1,57 |
|||||||||||
|
Желтый скорпион Anomalobuthus rickmersi |
2 |
2 |
0,52 |
|||||||||||
|
Саранчи род Calliptamus |
8 |
11 |
1 |
1 |
21 |
5,5 |
||||||||
|
Медведка обыкновенная Gryllotalpa unispina |
4 |
1 |
5 |
1,31 |
||||||||||
|
Афодии Aphodius sp. |
1 |
1 |
1 |
3 |
0,79 |
|||||||||
|
Медляки Blaps sp. |
2 |
2 |
0,52 |
|||||||||||
|
Чернотелок Adesmia sp. |
3 |
3 |
0,79 |
|||||||||||
|
Чернотелок Pisterotarsa sp. |
2 |
2 |
0,52 |
|||||||||||
|
Чернотелок Cyphogenia sp. |
1 |
1 |
0,26 |
|||||||||||
|
Жужелицы-бегун Ophonus sp. |
4 |
1 |
5 |
1,31 |
||||||||||
|
Златка пятнистая Julodis variolaris |
6 |
6 |
1,57 |
|||||||||||
|
Всего беспозвоночных |
8 |
24 |
13 |
9 |
2 |
56 |
14,66 |
|||||||
|
Рептилии |
||||||||||||||
|
Степная агама Trapelus sanguinolentus |
3 |
1 |
4 |
1,05 |
||||||||||
|
Удавчик восточный Eryx tataricus |
2 |
2 |
0,52 |
|||||||||||
|
Всего рептилий |
5 |
1 |
6 |
1,57 |
||||||||||
|
Птицы |
||||||||||||||
|
Чомга Podiceps cristatus |
1 |
1 |
0,26 |
|||||||||||
|
Желтая цапля Ardeola ralloides |
1 |
1 |
0,26 |
|||||||||||
|
Чирок-свистунок Anas crecca |
2 |
2 |
0,52 |
|||||||||||
|
Обыкновенный перепел Coturnix coturnix |
1 |
1 |
0,26 |
|||||||||||
|
Лысуха Fulica atra |
2 |
2 |
0,52 |
|||||||||||
|
В размер постушок* |
1 |
1 |
0,26 |
|||||||||||
|
Саджа Syrrhaptes paradoxus |
1 |
1 |
2 |
0,52 |
||||||||||
|
Чернобрюхий рябок Pterocles orientalis |
1 |
1 |
0,26 |
|||||||||||
|
Хохлатый жаворонок Galerida cristata |
1 |
1 |
0,26 |
|||||||||||
|
Черная ворона Corvus corone |
1 |
1 |
0,26 |
|||||||||||
|
Всего птиц |
1 |
10 |
2 |
13 |
3,4 |
|||||||||
|
Млекопитающие |
||||||||||||||
|
Еж ушастый Hemiechinus auritus |
3 |
3 |
6 |
1,57 |
||||||||||
|
Малая белозубка Crocidura suaveolens |
1 |
1 |
2 |
0,52 |
||||||||||
|
Заяц-толай Lepus tolai |
1 |
5 |
17 |
4 |
27 |
7,07 |
||||||||
|
Пустынный кожан Eptesicus bottae |
1 |
1 |
0,26 |
|||||||||||
|
Поздний кожан Eptesicus serotinus |
1 |
1 |
2 |
0,52 |
||||||||||
|
Тушканчик малый Scarturus (Allactaga) elater |
1 |
2 |
1 |
2 |
1 |
7 |
1,83 |
|||||||
|
Песчанка гребенщиковая Meriones tamariscinus |
1 |
17 |
18 |
4,71 |
||||||||||
|
Песчанка краснохвостая Meriones libycus |
11 |
28 |
43 |
15 |
97 |
25,39 |
||||||||
|
Песчанка полуденная Meriones meridianus |
1 |
13 |
13 |
22 |
6 |
55 |
14,40 |
|||||||
|
Хомячок серый Nothocricetulus (Сricetulus) migratоrius |
8 |
4 |
7 |
19 |
4,97 |
|||||||||
|
Мышь домовая Mus musculus |
5 |
15 |
50 |
3 |
73 |
19,11 |
||||||||
|
Всего млекопитающих |
4 |
42 |
69 |
163 |
29 |
307 |
80,37 |
|||||||
|
Всего |
12 |
66 |
83 |
187 |
34 |
382 |
100 |
|||||||
Примечания: * – вид не определен.
Составлена авторами на основе полученных данных в ходе исследования.
На равнинах было найдено только 8 видов млекопитающих, 2 вида птиц, 1 вид рептилий и 4 вида беспозвоночных, всего 15 видов, то есть в 2 раза меньше видов, чем в зоне чинков. Важно отметить, что только в зоне чинков обнаружены восточный удавчик и 2 вида рукокрылых, а также 9 видов птиц, не обнаруженных в погадках их равнинных биотопов.
По результатам анализа трофического спектра филина на территории пустыни Аралкум, выявлено 382 объекта добычи. В структуре рациона доминировали млекопитающие – 80,37 %. Существенную, но второстепенную долю составили беспозвоночные – 14,66 %. Птицы и рептилии отмечены эпизодически – 3,40 и 1,57 % соответственно (табл. 1).
Среди млекопитающих наибольший вклад в пищевой рацион вносили грызуны, прежде всего песчанки. Абсолютным доминантом выступила Meriones libycus (25,39 %). Высокая встречаемость также характерна для Mus musculus (19,11 %) и Meriones meridianus (14,40 %). Значимым компонентом добычи являлся Lepus tolai (7,07 %), а также Nothocricetulus (Сricetulus) migratorius (4,97 %) и Meriones tamariscinus – 18 экз. (4,71 %). Реже Scarturus (Allactaga) elater (1,83 %), Hemiechinus auritus (1,57 %), Crocidura suaveolens (0,52 %), Eptesicus serotinus (0,52 %) и Eptesicus bottae (0,26 %) (табл. 1). В целом преобладание мелких пустынных грызунов указывает на их ведущую роль в кормовой базе филина в условиях аридных ландшафтов пустыни Аралкум.
Рептилии в рационе представлены двумя видами: Trapelus sanguinolentus (1,05 %) и Eryx tataricus (0,52 %), что характеризует их как редкую, факультативную добычу. Птицы также формируют малую долю рациона и представлены преимущественно видами, связанными с водно-болотными и открытыми биотопами: Anas crecca (0,52 %), Fulica atra (0,52 %), Syrrhaptes paradoxus (0,52 %); остальные виды встречены единично (0,26 % каждый): Podiceps cristatus, Ardeola ralloides, Coturnix coturnix, Pterocles orientalis, Galerida cristata, Corvus corone.
В группе беспозвоночных наибольшая численность отмечена у саранчовых рода Calliptamus (5,50 %), что, вероятно, отражает их массовость и доступность. В сопутствующем комплексе добычи представлены Galeodes caspius (1,57 %), Julodis variolaris (1,57 %), Gryllotalpa unispina (1,31 %), Ophonus sp. (1,31 %), Aphodius sp. (0,79 %), Adesmia sp. (0,79 %), Anomalobuthus rickmersi (0,52 %), Blaps sp. (0,52 %), Pisterotarsa sp. (0,52 %) и Cyphogenia sp. (0,26 %) (табл. 3).
Анализ биотопической приуроченности видов-жертв показал, что более высокое видовое богатство характерно для чинков и прилегающих к ним территорий по сравнению с равнинными участками. Вероятно, это связано с большей неоднородностью рельефа и высоким разнообразием микроместообитаний, формирующих благоприятные условия для пребывания различных групп позвоночных и беспозвоночных животных. Чинковые участки отличаются мозаичностью ландшафта, наличием трещин, уступов, каменистых выходов и участков с различным типом растительности, что создает многочисленные укрытия, места отдыха, ночевки и размножения для животных. Кроме того, подобные биотопы обеспечивают более широкий спектр кормовых ресурсов и защитных условий, что способствует концентрации потенциальных объектов добычи филина и повышению общего уровня биоразнообразия.
Рацион филина в пределах обследованной территории Аралкума характеризуется выраженной миофагией со смещением в сторону грызунов. Основу трофической структуры составляют песчанки рода Meriones и облигатный синантроп домовая мышь, что отражает как высокую численность данных видов в аридных и антропогенных экосистемах, так и их доступность для хищника. Птицы и рептилии выступают в качестве эпизодической добычи, их суммарная доля не превышает 5 %, что указывает на второстепенное значение этих таксонов в формировании рациона хищника. Доля беспозвоночных достигает 14,66 %, что является закономерным для аридных территорий, особенно в условиях массового размножения саранчовых (Calliptamus).
Вместе с тем, несмотря на сравнительно высокую численную представленность беспозвоночных в составе погадок, их вклад в энергетический баланс филина, вероятно, существенно ниже по сравнению с грызунами. Это связано с меньшей калорийностью и биомассой единичной добычи. Таким образом, при количественном анализе рациона наблюдается заметная роль беспозвоночных, однако с точки зрения трофической энергетики доминирующее значение сохраняют мелкие млекопитающие.
Анализ погадок, собранных на острове Возрождения, продемонстрировал значительное видовое разнообразие птиц. В исследуемом материале были идентифицированы кости и перья водоплавающих и прибрежных видов, включая Podiceps cristatus, Ardeola ralloides, Anas crecca и Fulica atra. На территории, расположенной примерно в 7 км от гнезда ночного хищника, находится единственный искусственный источник воды (скважина), что способствует концентрации большого количества птиц, особенно в период миграции (рис. 1). Птицы используют данный участок для кратковременного отдыха, кормления и ночевки, после чего вновь покидают его. На основании этих наблюдений можно заключить, что филин, вероятно, посещает этот участок и проявляет высокую эффективность трофической активности в отношении прилетающих птиц.
Ряд видов, зафиксированных в рационе филина на плато Устюрт, откуда гипотетически могло идти заселение бывших островов Аральского моря, не был обнаружен в составе погадок, собранных на территории Аралкума. Анализ данных, полученных на плато Устюрт [16; 17], показал, что большинство видов, идентифицированных в качестве добычи филина (H. auritus, L. tolai, S. elater, N. migratorius, M. libycus, M. meridianus, M. musculus), также встречались на исследуемой территории. Вместе с тем такие виды, как Diplomesodon pulchellum, Spermophilus fulvus, Allactaga major, A. jaculus, A. severtzovi, Ellobius talpinus, Rhombomys opimus, Nesokia indica и Mustela nivalis, в погадках филина Аралкума не были зафиксированы. Аналогичная тенденция наблюдалась и среди беспозвоночных: состав погадок плато Устюрт характеризовался более широким видовым разнообразием таксонов. Сравнительный анализ с данными более ранних исследований [11] показал, что отмеченный ранее Spermophilus fulvus, составлявший 2,7 % в рационе филина, в наших исследованиях не отмечен. В то же время виды E. bottae и E. serotinus, идентифицированные в настоящем исследовании, были впервые зафиксированы на территории Аралкума в качестве добычи филина, что расширяет сведения о трофической структуре его рациона в данном регионе.
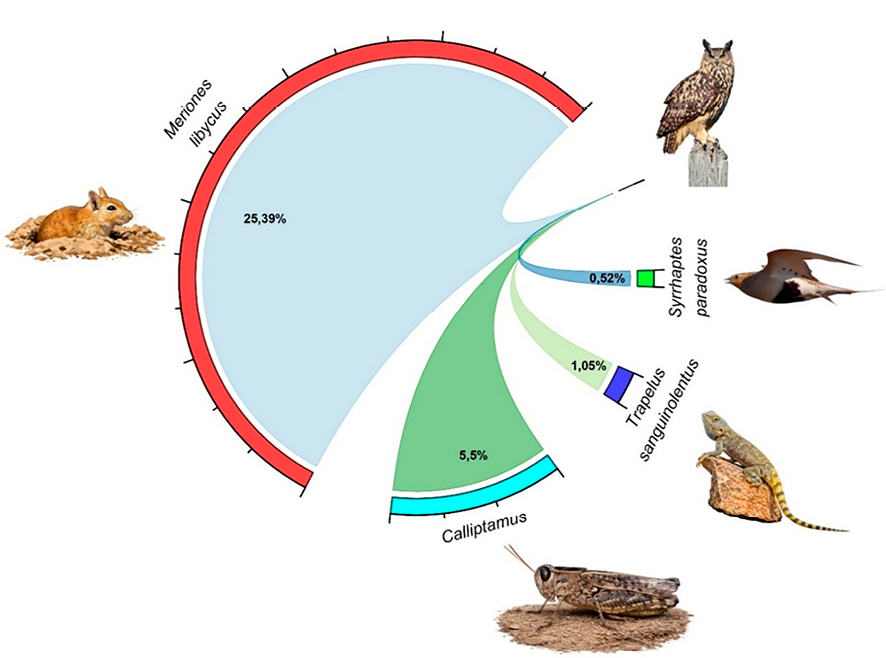
Рис. 2. Доминирующие виды добычи филина на территории Аралкума Примечание: составлен авторами по результатам данного исследования
При анализе рациона в разрезе крупных таксономических групп установлено, что среди беспозвоночных наибольшую долю составили представители рода Calliptamus (5,5%). В группе пресмыкающихся отмечен Trapelus sanguinolentus (1,05%). Среди птиц зафиксирован Syrrhaptes paradoxus (0,52%). Наибольший вклад в структуру рациона среди млекопитающих, как отмечалось ранее, внес Meriones libycus – 25,39 %, что свидетельствует о его доминирующем значении в питании исследуемого ночного хищника (рис. 2).
Несмотря на сравнительно невысокое видовое разнообразие птиц в составе добычи, по частоте встречаемости и территориальному распространению отчетливо выделяется саджа. Данный вид широко распространен в пределах исследуемого региона, регулярно регистрируется в полевых наблюдениях и играет заметную экологическую роль в пустынных экосистемах, что отражается и в его присутствии в трофическом спектре хищной птицы.
Сопоставление полученных данных с результатами анализа погадок филина, собранных на территории плато Устюрт, выявляет отчетливые различия как в видовом разнообразии добычи, так и в ее количественном распределении в составе рациона. Трофический спектр хищных птиц, обитающих в Устюртском регионе, отличается большей широтой по сравнению с Аралкумом, что, вероятно, обусловлено неоднородностью ландшафтов и более стабильными условиями обитания потенциальных кормовых объектов плато Устюрт по сравнению с обедненной фауной островного типа Аралкума, находящейся в процессе трансформации.
Заключение
Выявленные различия, по-видимому, отражают региональную специфику фауны, различия в структуре биотопов и доступности кормовых ресурсов, а также особенности формирования молодой пустынной экосистемы Аралкума по сравнению с более устойчивыми природными комплексами плато Устюрт.
Таким образом, анализ пищевого рациона показал, что он связан как с особенностями биoтопических условий пребывания видов жертв, так и с биологическими особенностями хищника. Трофическая структура филина на территории пустыни Аралкум характеризуется выраженным доминированием млекопитающих (преимущественно грызунов) при подчиненной роли беспозвоночных и эпизодическом участии птиц и рептилий.
Библиографическая ссылка
Ёркулов Ж. М., Быкова Е. А., Есипов А. В., Умаров С. С., Ражабова Д. А. ОСОБЕННОСТИ ПИТАНИЯ ФИЛИНА (BUBO BUBO) В ТРАНСФОРМИРОВАННЫХ ЭКОСИСТЕМАХ АРАЛКУМА // Научное обозрение. Биологические науки. 2026. № 3. С. 22-30;URL: https://science-biology.ru/en/article/view?id=1463 (дата обращения: 03.07.2026).