

 science-review.ru
science-review.ru
Алтайский край занимает юго-восточную часть Западной Сибири, располагается в бассейне верхнего течения р. Оби. Территория края отличается большим разнообразием рельефа: от равнин Кулунды, Приобского плато, Бие-Чумышской возвышенности и до Предалтайской и Предсалаирской предгорных равнин, а также гор Салаирского кряжа и части Русского Алтая. Соответственно рельефу по территории региона в определенной степени распределяется видовое и популяционное разнообразие растительного мира [1]. Алтайский край принадлежит к числу самых развитых сельскохозяйственных регионов России, кроме того, на его территории активно расширяется туристско–рекреационная сфера, что негативно сказывается на биоразнообразии. Несмотря на это, даже в таких условиях природа края сохраняет богатое растительное разнообразие. Флора Алтайского края представлена более 2100 видами высших сосудистых растений, в том числе – свыше 1800 аборигенных и 300 адвентивных, около 400 видов мхов, около 700 видов лишайников [2].
Значительная часть флоры произрастает на территории Бие-Чумышской возвышенности, которая представляет собой увалисто-волнистую равнину, расположенную в возвышенной лесостепи (275–400 м). Территория возвышенности в значительной степени расчленена овражно-балочной сетью, имеют место отрицательные элементы рельефа: долины рек, логов и положительные – увалы, что расширяет диапазон экологических параметров местообитания растений [3–5]. За многие годы деятельность человека привела к преобразованию исходной флоры Бие-Чумышской возвышенности в антропогенно-трансформированную, возросла степень редкости в нарушенных экосистемах представителей многих видов растений, численность их ценопопуляций сокращается.
Сложившееся в обществе мировоззрение не позволяет изменить сегодняшний стереотип природопользования на Алтае. Достоверные факты экологического неблагополучия окружающей природной среды затрудняют понимание сущности современных природоохранных задач [6–8]. Становятся все более актуальными вопросы изучения растений в природной обстановке, подверженных возрастающему негативному воздействию и снижающих свою численность.
Семейство Orchidaceae Juss. – одно из крупнейших среди покрытосеменных, насчитывает около 25 тыс. видов [9, 10] и занимает заслуженное место среди растений, требующих особого внимания, связанного с их биологией и экологией, отдельные вопросы изучения которых далеки от завершения [11–14]. Практически все виды семейства орхидных являются редкими, об этом говорит тот факт, что численность известных в природе популяций сокращается по всему миру [15, 16]. Более половины представителей семейства орхидных на территории нашей страны внесены в Красные книги федерального и региональных уровней [17]. Общая численность видов семейства орхидных на территории Алтайского края – 27, из них 10 видов внесены в Красные книги [18].
В качестве объекта исследований нами выбрана ценопопуляция Gymnadenia conopsea (L.) R. Br. (Orchidaceae), расположенная в Бие-Чумышской возвышенности в верховье бассейна реки Ангуреп (окрестности с. Овсянниково Целинного района Алтайского края).
Цель настоящей работы – выявить местонахождение и численность ценопопуляции G. conopsea, изучить отдельные морфометрические параметры, сопутствующие виды, лимитирующие факторы и предложить первоочередные меры охраны.
Материалы и методы исследования
В основу работы положены данные, полученные авторами в полевых экспедициях, при этом использовались отдельные методы популяционной биологии растений [19, 20], популяционного мониторинга редких и исчезающих видов растений [21], методы полевых экологических исследований [22], а также сравнительно-географический и картографический методы. Особенность размещения особей в ценопопуляции оценивалась визуально. Выделен диффузно-рассеянный тип пространственной структуры, при котором особи рассеяны небольшими группами [23]. По числу особей исследованная ценопопуляция, согласно Т.М. Быченко [21], относится к малочисленной группе (до 100 особей).
Результаты исследования и их обсуждение
Кокушник длиннорогий – Gymnadenia conopsea (L.) R. Br. клубнекорневое многолетнее растение высотой 35–60 см [24, 25]. Клубни пальчато-лопастные. Стебель при основании одет буроватыми чешуевидными линейно-ланцетными листьями в количестве от 5 до 7. Листья слегка сложенные вдоль средней жилки, на концах стянутые в «башлычок». Соцветие – цилиндрический колос 7–18 см, густое из мелких розово-фиолетовых цветков (рис. 1).

Рис. 1. Кокушник длиннорогий. Фото С.В. Важова (01.07.2017)
Околоцветник простой, венчиковидный, неправильный. Губа трехлопастная, с нитевидным серповидно изогнутым в виде бараньего рога шпорцем, в 1,5–2 раза длиннее завязи. Предположительно, что эта особенность дала основание для видового названия растения [26]. Цветки выделяют обильный ароматный нектар, который принято называть гвоздичным. Аромат цветков усиливается к концу дня, что привлекает разнообразных насекомых-опылителей. В опыление кокушника длиннорогого основной вклад вносят медоносные и дикие пчелы, шмели, значительно участие ночных бабочек и жуков [26, 27]. Плод – удлиненная коробочка.
Биолого-экологические особенности. G. conopsea цветет в июне-июле, плодоносит в августе-сентябре, размножается семенами, изредка отмечается вегетативное размножение. Более половины цветков формируют плоды, которые, попадая на почву, прорастают. В течение 2-х лет проросток находится под землей и только на 3-й год жизни прорастает и появляется на поверхности почвы в виде зеленого листа. При благоприятных условиях роста и развития G. conopsea зацветает на 6–7 год [26]. Такой длительный период, предшествующий цветению кокушника длиннорогого, по сравнению с другими семействами, в определенной степени объясняет малочисленность цветущих групп этой орхидеи в селитебных ландшафтах.
G. conopsea произрастает на равнине и в условиях горного рельефа достигает альпийского пояса [24]. Предпочитает широкий диапазон водного режима почвы – от сухо-степного до болотного типа увлажнения, произрастает на лугах, опушках смешанных и мелколиственных лесов, по берегам рек. Растет на слабокислых или слабощелочных почвах. В Алтайском крае произрастает в Бийском, Волчихинском, Змеиногорском, Курьинском, Панкрушихинском, Солонешенском и Чарышском районах [28].
Ценопопуляция кокушника длиннорогого – G. conopsea изучена нами в 2017 г. в окрестностях с. Овсянниково Целинного района Алтайского края. Популяция вида занимает общую площадь 4 × 8 = 32 м2, состоит из 4-х групп скопления, способ размещения особей в группах – диффузно-рассеянный (табл. 1).
Изменение численности растений в группах скопления ценопопуляции G. conopsea проявляется в возрастной структуре и других характеристиках. Сокращение особей, сохранение или уменьшение генеративных частей групп, преобладание в онтогенетическом спектре вегетативных растений обусловлено эколого-ценотическими показателями местообитания [21, 29]. Общее количество растений в 4-х исследованных группах изменялось от 8 до 14, в том числе генеративных – от 1 до 4, вегетативных – от 7 до 10 (рис. 2).
В связи с условиями произрастания в отношении типов онтогенетических спектров ценопопуляции можно сделать вывод о том, что для склона холма юго-западной экспозиции сухо-степного типа увлажнения в Бие-Чумышской возвышенности характерны в основном молодые особи (70–88 %). Молодая возрастная структура ценопопуляции при значительном количестве молодых особей способствует снижению негативных последствий влияния неблагоприятных погодных условий на численность популяции. Наличие небольшого количества генеративных особей (12–30 %) обеспечивает достаточное поступление семян, а способность растений в неблагоприятных условиях переходить к семенной репродукции уменьшает долю взрослых вегетативных особей в онтогенетическом спектре [21, 29]. Морфометрические параметры генеративных особей кокушника длиннорогого варьировали в значительных пределах: высота побега – 35–59 см, количество листьев – 5–7 шт/раст., длина листа – 8–17 см, ширина листа – 0,5–2,0 см, длина соцветия – 7–18 см, количество цветков на побеге – 35–51 шт.
Как видно из табл. 2, в 1-й группе высота растений изменялась от 28,3 до 41,9 см (в среднем 35,3 ± 1,77 см), количество цветков на побеге – от 26,8 до 45,7 шт. (в среднем 36,9 ± 2,21 шт.). Полученные результаты в 1-й группе несколько отличаются от показателей других групп, но динамику их данных можно назвать однотипной.
Изучение сопутствующих видов в фитоценозе показало, что совместно с кокушником длиннорогим присутствуют 14 основных видов из 11 семейств (Poaceae, Scrophulariaceae, Rubiaceae, Plantaginaceae, Caryophyllaceae, Lamiaceae, Clusiaceae, Rosaceae, Fabaceae, Convallariaceae, Apiaceae (табл. 3).
Таблица 1
Характеристика местообитания ценопопуляции Gymnadenia conopsea (L.) R. Br.
|
№ группы |
Местообитание |
Количество генеративных особей |
Фенофаза |
Количество растений |
|
1 |
Склон холма юго-западной экспозиции сухо-степного типа увлажнения |
3 |
Цветение |
10 |
|
2 |
4 |
Цветение |
14 |
|
|
3 |
1 |
Цветение |
8 |
|
|
4 |
2 |
Цветение |
11 |
Примечание. Координаты: 52 °49'36,3" с.ш.; 86 °06'12,7" в.д.; абс. высота 360 м.
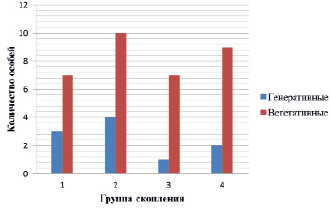
Рис. 2. Соотношение генеративных и вегетативных молодых особей G. conopsea по группам скопления
Таблица 2
Морфометрические параметры генеративных особей Gymnadenia conopsea (L.) R. Br.
|
Показатель |
Высота, см |
Количество цветков на побеге, шт. |
||||||
|
группа скопления |
группа скопления |
|||||||
|
1 |
2 |
3 |
4 |
1 |
2 |
3 |
4 |
|
|
M |
35,3 |
44,9 |
47,1 |
55,6 |
36,9 |
45,4 |
38,8 |
48,9 |
|
m |
1,77 |
1,86 |
2,07 |
3,19 |
2,21 |
2,01 |
1,35 |
1,78 |
|
min |
28,3 |
36,5 |
34,7 |
39,7 |
26,8 |
38,5 |
34,6 |
39,9 |
|
max |
41,9 |
50,7 |
55,9 |
63,9 |
45,7 |
55,7 |
46,7 |
54,4 |
Примечание. M – среднее арифметическое; m – стандартная ошибка; min/max – минимальное/максимальное значение признака.
Таблица 3
Сопутствующие виды в ценопопуляции Gymnadenia conopsea (L.) R. Br.
|
№ п/п |
Вид |
Шкала, балл |
|
|
О. Друде |
Ж. Браун-Бланке |
||
|
1 |
Ковыль перистый – Stipa pennata L. |
Cop3 |
4 |
|
2 |
Мытник – Pedicularis sibirica Vved. |
Cop2 |
3 |
|
3 |
Подмаренник – Galium ruthenicum Willd. |
Cop1 |
2 |
|
4 |
Подорожник Урвиллея – Plantago urvillei Opiz. |
Cop1 |
2 |
|
5 |
Вероника седая, беловойлочная – Veronica incana L. |
Sp |
1 |
|
6 |
Гвоздика разноцветная, степная – Dianthus versicolor Fisch. ex Link |
Sp |
1 |
|
7 |
Душица обыкновенная – Origanum vulgare L. |
Sp |
1 |
|
8 |
Зверобой продырявленный – Hypericum perforatum L. |
Sp |
1 |
|
9 |
Кизильник черноплодный – Cotoneaster melanocarpus Fisch. ex Blytt |
Sp |
1 |
|
10 |
Клевер люпиновидный – Trifolium lupinaster L. |
Sp |
1 |
|
11 |
Купена душистая – Poligonatum odoratum Mill. |
Sp |
1 |
|
12 |
Люцерна серповидная – Medicago falcata L. |
Sp |
1 |
|
13 |
Пустореберник обнаженный – Cenolophium denudatum (Hornem.) Tutin (C. fischeri (Spreng.) Koch) |
Sp |
1 |
|
14 |
Чина луговая – Lathyrus pratensis L. |
Sp |
1 |
Отдельные из сопутствующих видов более активно заселяют открытые пространства территории и в экологическом аспекте составляют реальную конкуренцию G. conopsea. Оценка по обилию по О. Друде и по обилию – покрытию по Ж. Браун-Бланке [21] показала, что Stipa pennata встречается очень обильно, но сплошного смыкания надземных частей не происходит, расстояние между особями 12–17 см, покрытие 60–65 %; Pedicularis sibirica Vved. встречается обильно, расстояние между особями 26–37 см; покрытие 40–45 %; Galium ruthenicum Willd. и Plantago urvillei Opiz. встречаются довольно обильно, расстояние между растениями – 57–82 см, покрытие не превышает 20 %; остальные 10 видов размещены в фитоценозе редко, рассеянно, в небольшом количестве, расстояние между особями – 95–120 см, покрытие менее 5 %.
Лимитирующие факторы. Кокушник неустойчив к сбору соцветий на букеты; слабо устойчив к сенокошению во время цветения и плодоношения, вытаптыванию животными, перевыпасу и пожарам; губительными для орхидеи являются культуртехнические и осушительные мелиорации; в последние годы подвержен негативному влиянию нерегулируемой рекреации [17, 26, 30].
Для условий Алтайского края данных по биологии, численности и степени воздействия хозяйственной деятельности на состояние ценопопуляций G. conopsea в настоящее время крайне мало. Однако именно они являются основой для совершенствования научно обоснованных рекомендаций по охране G. conopsea. Необходим систематический мониторинг за состоянием природных популяций G. conopsea, так как кокушник может быть сохранен только в естественных условиях. Также необходимы наблюдения за численностью на территории края, особенно в селитебных ландшафтах, запрет сбора растений и реальный контроль за его выполнением, совершенствование имеющихся и разработка новых технологий окультуривания вида, учитывающих сложность симбиоза орхидеи со специфическими грибами, особенно в первые годы жизни.
Заключение
Кокушник длиннорогий – Gymnadenia conopsea (L.) R. Br., как лекарственное, декоративное и медоносное растение, нуждается в охране. Этот вид предрасположен к снижению численности в связи с расширением аграрного и рекреационного освоения территории региона. В изученном местообитании орхидея произрастает в сравнительно благоприятных условиях. Структура ценопопуляции не зависит от географических особенностей территории. Биоморфологические признаки характеризуются хорошими показателями, однако отмечаются естественные факторы сокращения численности популяции путем вытеснения другими видами, механизм которого требует дальнейших исследований.
Данные, приведенные в статье, получены при выполнении темы НИР: «Реализация краеведческого подхода в географическом образовании через научно-исследовательскую деятельность», номер госрегистрации № АААА-А17-117011000005-0.
Библиографическая ссылка
Важов С.В., Важов В.М., Бахтин Р.Ф., Важова Т.И. ЦЕНОПОПУЛЯЦИЯ GYMNADENIA CONOPSEA (L.) R. BR. (ORCHIDACEAE) В АЛТАЙСКОМ КРАЕ // Научное обозрение. Биологические науки. 2017. № 4. С. 5-10;URL: https://science-biology.ru/ru/article/view?id=1076 (дата обращения: 01.07.2026).