

 science-review.ru
science-review.ru
Идея конкурентного сосуществования деревьев пронизывает множество исследований лесных насаждений. Имеются попытки моделирования развития одноярусных древостоев [1, 2], а также моделей сложных [3] и простых [4] насаждений, где конкуренция за ресурсы питания принята в качестве главного биотического фактора, со сложными расчетами пропорциональных площадей питания и областей доминирования. Конкурентная концепция древостоя вытекает из давних представлений о том, что лесной фитоценоз является сообществом растений, организованных борьбой за существование. Однако есть предельно старые леса, где конкуренция между деревьями должна закончиться, так как древостой образует устойчивые социальные структуры (биогруппы), где пики минерального питания и прирост разнесены во времени на 1–3 недели [5]. Генезис таких социальных структур начинается в самом раннем возрасте, и они сохраняются вплоть до спелости насаждений [1, 5]. Это позволяет рассматривать деревья как организмы, питание которых разнесено во времени и в пространстве, а значит, к ним применимы правила и законы экологии, в том числе концепция экологических ниш Мак-Артура [6].
Обсуждение применяемых для этого методов и первые результаты по изучению роста сосны в зависимости от расстояний до соседних деревьев в старом сосновом насаждении показало, что расстояние до ближнего соседа весьма слабо влияет на диаметр соседствующих деревьев при r = 0,148 ± 0,063. При этом сила влияния этого расстояния на их средний диаметр составляет всего лишь 4 % [7]. Эти данные позволяют предполагать, что в старых насаждениях конкурентное давление за ресурсы питания снижается до столь незначительных величин, что ими уже можно пренебречь в моделях структуры таких предельно старых древостоев.
Цель данной работы: выяснить структуру расположения деревьев в старом сосновом древостое и определить влияние взаимодействия (конкуренции) между деревьями на их размеры. В частности, влияет ли размер (рост) дерева на размер другого дерева, соседнего с ним? Если размер второго дерева снижается при увеличении размера первого, то это будут конкурентные отношения с подавлением роста. Однако если второе дерево точно так же увеличивает свои размеры при увеличении размеров первого, то это будут уже отношения партнерства, но никак не конкуренции в обычном ее понимании, когда один из партнеров рано или поздно начинает доминировать, а второй снижает свои размеры. Это упрощенное понимание, но оно позволяет заострить вопрос о том, в чем же собственно проявляется конкуренция, и можно ли измерить ее результат.
Объект работ – массив сосны обыкновенной (Рinus sylvestris L.) в Пермском городском лесничестве (леса г. Перми), в кв. 61 Нижне-Курьинского участкового лесничества, где на площади 2 га сохранились деревья с диаметрами до 68 см. Древостой оставляет хорошее впечатление; его структура не нарушена, так как вывозку древесины от санитарных рубок осуществляли по просветам между деревьями. Были заложены две пробные площади: участок «Южный» и участок «Северный» площадью 1,09 и 1,11 га соответственно, примыкающие друг к другу. Каждое дерево, всего 736 шт., нанесли на план в масштабе 1:100 с точностью ± 10–35 см. Взаимное расположение деревьев в биогруппах определялось точнее (± 3–5 см). Для работы карта была оцифрована в программе ArcMap. У каждого дерева делали отметку на высоте 1,3 м, где измеряли окружность ствола с округлением до 0,5 см, после чего расчетным путем определяли диаметр с округлением до ± 0,1 см. Возраст определили в 2017–2018 гг. по кернам древесины на высоте 1,3 м. Для определения высоты яруса сосны измеряли 149 деревьев. История заселения массива сосной связана со сплошной рубкой леса и выжиганием угля на этой территории в 1820-е гг. В 2003 г. (14 лет назад) здесь прошел ветровал с повреждением около 7 % деревьев.
Таксационная характеристика насаждений на участках в 2017 г. оказалась следующей.
Участок «Южный» – формула состава 10СедЕ, возраст 184 года, средняя высота 32,1 м, средний диаметр 39,8 см, тип леса сосняк кисличниковый с преобладанием 70 % черники и 30 % брусники, класс бонитета 1, абсолютная полнота 42,8 м2/га, относительная полнота 0,91, запас 614 м3/га. Если учесть 29 ветровальных деревьев, то по отношению к ныне имеющимся (376 шт.) их число и, соответственно, полнота и запас могли бы увеличиться на 7,7 % и составить 660 м3/га при полноте 0,98. Текущая густота 344 шт/га.
Участок «Северный» имеет тот же состав, возраст и близкий тип леса – сосняк кисличниковый, но здесь уже явно преобладает черника (95 %), что свидетельствует о несколько более влажных условиях. Средняя высота 33,5 м, средний диаметр 42,1 см. Густота древостоя меньше на 6 %, но полнота выше и достигает 0,95, запас составляет 655 м3/га. Если же учесть 23 ветровальных дерева, что составляет 6,5 % к ныне живым деревьям, то по отношению к ним (359 шт.) их число, полнота и запас древостоя могли бы увеличиться на 6,5 % и составить 700 м3/га при полноте 1,01. Текущая густота здесь 324 шт/га.
На обоих участках единично встречается ель в возрасте 120 лет с диаметрами 20–45 см, образующая невыраженный второй ярус. Представлен также подрост ели в основном поколением высотой 3–6 м в возрасте 35 лет, возникшим после низового пожара и двойного обильного урожая семян ели в 1981 и 1982 г. Сравнение данных таксации показало достоверное увеличение среднего диаметра сосны на участке «Северный» на 5,5 % и высоты на 3 %. В целом данные таксации подтвердили предположения о том, что на участке «Северный» условия более благоприятны. Это могут быть как лучшие почвенные условия, так и меньшая на 6 % густота, при которой сформировалось больше крупных деревьев.
Результаты исследования и их обсуждение
Самое короткое из ближних расстояний между деревьями составило 84 см, а самый дальний «ближний» сосед был обнаружен на расстоянии 790 см. Всего на участках «Южный» и «Северный» было учтено 244 и 235 расстояний соответственно. Их частоты показаны на диаграммах, из которых становится понятно, что на участке «Северный» соседи располагаются заметно дальше (в среднем 350 см), чем на участке «Южный» (в среднем 315 см), что обусловлено меньшей густотой участка «Северный». Модальные классы расстояний на этих участках также различаются на 100 см (рис. 1).


Рис. 1. Расстояния между ближними соседствующими деревьями в 184-летнем сосновом древостое на участках «Северный» и «Южный»
Таблица 1
Соседствующие деревья и корреляция (r) диаметра более крупного дерева с его малым соседом
|
Расстояние между соседями, см |
Участок «Южный» |
Участок «Северный» |
Оба участка |
||||||
|
n, шт. |
r |
ошибка r |
n, шт. |
r |
ошибка r |
n, шт. |
r |
ошибка r |
|
|
84–170 |
27 |
0,31* |
0,17 |
24 |
0,67 |
0,11 |
51 |
0,48 |
0,11 |
|
171–270 |
67 |
0,53 |
0,09 |
45 |
0,55 |
0,10 |
112 |
0,54 |
0,07 |
|
271–350 |
65 |
0,26 |
0,12 |
56 |
0,67 |
0,07 |
121 |
0,45 |
0,07 |
|
351–450 |
57 |
0,46 |
0,10 |
57 |
0,54 |
0,09 |
114 |
0,50 |
0,07 |
|
451–800 |
28 |
0,34 |
0,17 |
53 |
0,23* |
0,13 |
81 |
0,27 |
0,10 |
|
Общие |
244 |
0,429 |
0,052 |
235 |
0,509 |
0,048 |
479 |
0,463 |
0,036 |
|
В том числе |
|||||||||
|
для 84–450 см |
216 |
0,441 |
0,055 |
182 |
0,589 |
0,048 |
398 |
0,520 |
0,037 |
|
0,31* – корреляция недостоверна |
|||||||||
Далее эти две совокупности были разбиты на пять выборок. За основу было взято расстояние в 270 см, которое составляет ½ от среднего расстояния между деревьями на обоих участках, составляющего 5,45 м, и которое послужило основанием для градуировки взаимодействия и отнесения соседствующих деревьев к биогруппе. Это расстояние разделили на две части – до 170 см и от 171 до 270 см. Оставшийся промежуток 271–800 см разделили на три неравных интервала с расчетом, чтобы в них оказались выборки, достаточные по объему для определения корреляций. Корреляции рассчитывали после ранжирования диаметров в парах, т.е. на первое место ставили дерево с большим, а на второе – с малым диаметром. В результате получили следующие данные (табл. 1).
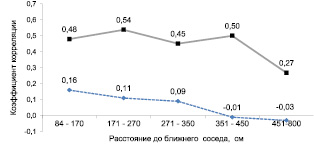
Рис. 2. Корреляции между диаметрами крупного и малого дерева у соседствующих деревьев (вверху) и у случайных пар деревьев (внизу)
Сравнение общих корреляций показывает их повышение от r = 0,429 на участке «Южный» и до r = 0,509 на участке «Северный». Это повышение, однако, недостоверно даже при t =1,12 < t0.10 = 1,66, и поэтому корреляции по этим участкам можно объединить. Полученные коэффициенты корреляций диаметра крупного дерева с его малым соседом оказались достоверно выше корреляций в контроле, рассчитанных в случайных парах, также образованных крупным и малым деревом (рис. 2).
Поэтому далее мы взяли объединенный диапазон расстояний от 84 до 450 см, исключая дальние расстояния. Полученная при этом общая для обоих участков корреляция оказалась равна r = 0,52, а ее отличие с корреляцией на расстояниях 451–800 см, равной r = 0,27, оказалось достоверно при t = 2.33 > t0.05 = 2.0. В этом диапазоне расстояний получились достоверные отличия в корреляциях также и между «Южным» и «Северным» участками, где они оказались равны соответственно, r = 0,441 ± 0,055 и r = 0,509 ± 0,048 при t = 2,02 > t0,05 = 1,98. Участок «Южный» имеет несколько большую сухость почвы и большую текущую густоту, и поэтому снижение корреляций на нем вполне может объясняться действием этих факторов. Однако утверждать это однозначно пока нельзя. Данное явление, насколько нам стало известно после обзора литературы и анализа моделей динамики роста и развития древостоев [1] изучается вообще впервые. Данных пока мало, и различие по взаимным корреляциям размеров деревьев-соседей по экотопам может быть только на этих двух конкретных участках.
Однако взаимодействие между деревьями может быть рассмотрено и в совершенно иной плоскости – в концепции экологической ниши, которую деревья делят между собой десятилетиями. И вопрос в таком контексте также рассматривается впервые.
Важно внимательно рассмотреть экспериментальные факты с показателями размеров деревьев в биогруппах и в контроле, т.е. в парах деревьев, которые были образованы первым крупным и вторым более мелким деревом в биогруппе, а в контроле пары образовывали каждое дерево из биогруппы с ближайшим деревом за ее пределами (табл. 2).
Таблица 2
Отношение максимального диаметра дерева в паре (Дмах) к минимальному (Дмin) в биогруппах и в контроле на участке «Южный»
|
Показатели |
Статистики |
|||||
|
n |
x |
± m |
СV, % |
min |
max |
|
|
Биогруппы из двух деревьев (56 шт.) |
||||||
|
Диаметр, см |
112 |
39,0 |
0,76 |
20,6 |
24 |
65 |
|
Расстояние, см |
56 |
195 |
7,0 |
26,8 |
84 |
271 |
|
Дмах /Дмin |
56 |
1,272 |
0,03 |
20,1 |
1,01 |
2,14 |
|
Контрольные пары деревьев-соседей (56 шт.) |
||||||
|
Диаметр, см |
112 |
39,2 |
0,70 |
18,9 |
25 |
63 |
|
Расстояние, см |
112 |
525 |
19,0 |
38,4 |
126 |
1427 |
|
Дмах /Дмin |
112 |
1,274 |
0,022 |
18,2 |
1,00 |
2,29 |
Полученное отношение Дmax / Дmin, при округлении одинаковое в биогруппах и контроле и равное 1,27, можно сравнить с так называемой «постоянной Хатчинсона». Эту постоянную предложил Дж. Хатчинсон в 1959 г. для оценки конкуренции среди видов птиц, животных и насекомых, занимающих разные экологические ниши в пределах одного ареала. Ее значение рассчитывается отношением массы или размеров тела более крупных видов к соседствующим мелким видам; она равна для массы тела 2,0 и для линейных размеров 1,3 при минимальной конкуренции видов за ресурсы питания [6].
На наш взгляд, идея ввода «постоянной Хатчинсона» была нужна для понимания того, что при таком ее значении конкуренция между близкими видами фауны за ресурсы питания минимальна, и они прекрасно сосуществуют вместе. По аналогии можно считать, что деревья-соседи – это разные организмы, имеющее различия по множеству показателей и по разному питающиеся (занимающие разные экологические ниши в освоении доставшихся им ресурсов питания). При округлении полученного нами Дмаx / Дмin получаем значение 1,27 ≈ 1,3. То есть данные «из фауны» Дж. Хатчинсона вполне адекватно совпадают с нашими данными «из флоры» по сосне. Это весьма неожиданно, и подтверждает глубинную связь биологических явлений, которые приходится нащупывать порой интуитивно.
Таким образом, изучение структуры расположения 736 деревьев сосны на двух участках 184-летнего насаждения 1 класса бонитета на общей площади 2,2 га на супесчаной почве, несколько отличающихся увлажнением (сосняк чернично-брусничниковый и сосняк кисличниковый), с полнотой 0,91 и 0,95 позволяет сделать следующие выводы.
Выводы
1. Живые деревья сосны растут в 184 года от ближнего соседа на расстояниях от 84 до 790 см; в 82 % случаев диапазон расстояний составляет 150–450 см.
2. Коэффициент корреляции диаметра более крупного дерева в биогруппе с его малым соседом достоверно выше корреляций в контроле, рассчитанных в случайных парах в той же выборке, также образованных крупным и малым деревом. При этом корреляция на расстояниях между соседями 84–450 см оказалась равна r = 0,52 ± 0,037, а с увеличением расстояний до 451–800 см она достоверно понижалась до r = 0,27 ± 0,10.
3. Достоверная связь между диаметрами пар деревьев в биогруппах указывает на явление, которое можно назвать «толерантностью» или «партнерством», в отличие от конкуренции, которая должна приводить, наоборот, к рассогласованию их роста.
4. Отношение максимального диаметра дерева в соседствующих парах к минимальному равно в среднем 1,3, что совпадает с «постоянной Хатчинсона» и характеризует древостой как сообщество, где к 184 годам конкуренция за ресурсы питания между деревьями становится минимальной, что может быть объяснено известным из литературы наступлением пиков ежегодного прироста (и пиков питания) с различиями в 1–2 недели.
Библиографическая ссылка
Рогозин М.В. ПОСТОЯННАЯ ХАТЧИНСОНА И КОНКУРЕНЦИЯ В СОСНОВОМ ДРЕВОСТОЕ // Научное обозрение. Биологические науки. 2019. № 1. С. 51-55;URL: https://science-biology.ru/ru/article/view?id=1134 (дата обращения: 27.06.2026).