

 science-review.ru
science-review.ru
Из множества аспектов исследования механизмов, обеспечивающих оптимальное функционирование центральной нервной системы, достаточно большой период времени активно изучаются структурно-функциональные возможности гематоэнцефалического барьера (ГЭБ) [1, 2].
Именно наличие ГЭБ позволяет поддерживать гомеостаз всего организма, в том числе и центральной нервной системы [3, 4]. Наиболее функционально значимой структурой признаны plexus choroids головного мозга (ССГМ) [5, 6]. Очевидно, что важным является онтогенетическое становление морфологического субстрата ССГМ [7]. Несомненно, важна морфофункциональная состоятельность ССГМ не только в условиях нормы, но с учетом окружающей среды [8, 9]. Формирование любой структуры организма хордовых, включая и млекопитающих, обусловлено также окружающей средой, в частности воздействием стресса [10].
Однако, несмотря на большое внимание исследователей к особенностям функционирования центральной нервной системы, частные вопросы структуры и функций plexus choroids оставляют достаточно много вопросов по формированию барьера, в частности, в онто- и филогенезе. Требуют детализации преобразования структурных компонентов plexus choroids в историческом аспекте. Отсутствует исследование рандомизации морфофункционального состояния plexus choroids боковых, 3-его и 4-го желудочков ликворной системы головного мозга. Совершенно очевидно, что это позволит раскрыть механизмы защиты от проникновения агрессивных препаратов, способных влиять на гомеостаз в центральной нервной системе.
Было проведено сравнительное изучение структурно-функционального становления plexus choroids головного мозга позвоночных животных боковых, 3-го и 4-го желудочков и выявление особенностей функционального созревания каждого.
В работе использованы методы: микроанатомические (Тотальный пленочный препарат сосудистых сплетений головного мозга мышей по Л.Г. Сентюровой и Р.А. Зумерову, авторское свидетельство № 1288536), гематоксилин и эозин, Ван–Гизон, Харт; толуидиновый синий, по Нахласу, по Гомори и Гленнеру; метод импрегнации по В.В. Куприянову. Проводилось определение высоты клеток хориоидного эпителия (в мкм), средний диаметр их ядер (в мкм), толщина соединительнотканной стромы (в мкм), средний диаметр отдельных звеньев микроциркуляторного русла (в мкм). Статистическую обработку полученных данных осуществляли на персональном компьютере с использованием пакета «Анализ данных» в рамках программы Microsoft Excelи BioStat 2008Professional 5.8.4.
Материалом для исследования служили ССГМ миноги (3), карповых (3) и осетровых рыб (3), земноводных (3), птиц (3) и беспородных белых крыс (25), мышей (25). Всего было детально изучены ССГМ 65 особей. При работе использовали устройство для фиксации мелких лабораторных животных в эксперименте [11].
Результаты исследования и их обсуждение
Обнаружено, что у миноги в переднем мозге plexus choroids представлены эпителиальной пластинкой. Отмечается расположение эпителия в несколько слоев. Кровеносные сосуды тесно соприкасаются с этим эпителием, что позволяет обеспечивать максимальный функциональный контакт. Сосуды заполнены элементами крови.
У карповых ССГМ кроме сплетения в желудочке продолговатого мозга, они располагаются и в промежуточном мозге. У сазана, каспийской воблы, толстолобика, plexus choroids заметно развиты лишь в 3-м и 4-м желудочках. Дупликатура видоизмененной эпендимы с одной стороны образована цилиндрическими клетками, с другой – плоской формы. Первые характеризует практически не окрашивающаяся цитоплазма и апикальное расположение ядра. Эти же взаимоотношения структур наблюдались и у взрослых особей. Вероятно, это сформировано под влиянием среды обитания и особенностями функционирования эпителиальных клеток ССГМ. А у осетра plexus choroids переднего мозга представлено дупликатурой видоизмененной эпендимы медиальной стенки желудочка. Эпителиоциты (хориоэпителиоциты) довольно однородной плоской формой. Сосуды встречаются редко. Соединительнотканная ткань не определяется. Plexus choroids 3-го и 4-го желудочков мозга были образованы сосудистыми клубочками с тонкой эндотелиальной выстилкой, обеспечивающей оптимальные условия для местного метаболизма. В то же время у севрюги мы не обнаружили plexus choroids в конечном мозге.
Достаточно позднее формирование элементов ГЭБ в классе рыб, возможно, определяется их функциональным значением. Об этом свидетельствует и тот факт, что форма их мало меняется, а только биометрические показатели.
У взрослых амфибий plexus choroids являются структурно оформленными образованиями. Эпителий, обращенный в полость желудочка, образован однослойными клетками кубической формы. Каждый эпителиоцит содержит по одному ядру, чаще округлой формы. В ядре прослеживается нежная сеточка гетерохроматина. Цитоплазма слегка базофильна. Толщина соединительнотканной стромы существенно увеличена по сравнению с филогенетически более ранними формами. В сплетениях всех желудочков наблюдается стаз элементов крови. Это указывает на существенную функциональную особенность ССГМ в процессах метаболизма у земноводных. По-видимому, медленный кровоток способствует более качественному обмену жизненно необходимыми веществами.
Сравнение plexus choroids головного мозга рептилий с земноводными позволяет утверждать, что они присутствуют также в боковых, 3-м и 4-м желудочках. Наибольшую площадь занимают сплетения в боковых и в 4-м желудочках. Строение аналогично таковому у амфибий. Хорошо выражена гроздевидная часть. Эпителиоциты преимущественно кубической формы располагается в один слой на базальной мембране. Следует отметить, что высота эпителиоцитов сравнима с аналогичными клетками амфибий. Каждая клетка содержит по одному ядру округлой формы и содержит довольно много гетерохроматина. Под базальной мембраной располагается рыхлая соединительная ткань. В её толще залегают довольно многочисленные кровеносные сосуды. Присутствуют фибробласты, наблюдаются единичные пигментоциты. Меньше всего соединительной ткани в сосудистом сплетении третьего желудочка.
Впервые у птиц закладку plexus choroids сосудистых сплетений головного мозга можно наблюдать на 2-й день пренатального развития. Первыми образуются сплетения 4-го желудочка в виде складки эпендимы с зародышевой мезенхимой. Только к концу первой недели начинается гистогенез plexus choroids боковых желудочков. Однако, эпителий псевдомногорядный. Ближе к вылуплению он становится однорядным однослойным. Как и у более ранних эволюционных форм каждый эпителиоцит (хориоэпителиоцит) имеет ядро чаще овальной формы. Можно наблюдать ядрышко и небольшое количество гетерохроматина. Цитоплазма оксифильна. Лишь к середине второй недели нами наблюдались plexus choroids 3-го желудочка. После вылупления в plexus choroids регистрировалось продолжение морфофункциональной дифференцировки, при сохранении заметной гетерохронии созревания ССГМ боковых, 3-го и 4-го желудочков.
В классе млекопитающих (крысы, мыши) сначала формируются ССГМ 4-го желудочка. В начале 3-й недели (15–17 день пренатального развития) появляются закладки ССГМ в боковых желудочках и 3-го желудочка. Далее гистогенез сосудистых сплетений продолжается уже в постнатальном периоде, когда в 3-м и 4-м желудочках появляются ворсинки, увеличивающие поверхность для оптимального обмена веществ. На пленочных препаратах показано, что формирование сосудистого русла продолжается вплоть до полного созревания организма. Сравнительный анализ структуры ССГМ желудочков позволяет сделать вывод, что наибольшую функциональную нагрузку несут ССГМ третьего и четвертого желудочков.
ССГМ у человека имеют значительно большие размеры и сложность строения. В 3-м и 4-м желудочках plexus choroids образовано инвагинацией однослойной крыши. А plexus choroids боковых желудочков являются производными медиальной части матрикса. Раньше других наблюдается закладка сосудистого сплетения 4-го желудочка. Очень рано ССГМ начинают формировать вторичные ворсины. В 7 недель эмбрионального развития сплетения боковых желудочков имеют лишь короткие первичные ворсины. И только на 8-й неделе появляются первые ветвления. Строма построена из мезенхимы, содержащей различные клеточные элементы. В плодном периоде к 15 неделям plexus choroids заполняют большую часть полости боковых желудочков. В этот период ядра эпителиоцитов (хориоэпителиоцитов) располагаются ближе к апикальному полюсу, что говорит о высокой степени метаболических процессов, в частности секреторных. После 16 недель ядра эпителиоцитов занимают обычное положение (ближе к основанию клетки). Наличие гломуса в сосудистых сплетениях бокового и 4-го желудочков, оставляет необходимость дальнейшего изучения plexus choroids, для более полного представления о роли ССГМ в обеспечении гомеостаза центральной нервной системы.
Сравнительное исследование филогенеза и отдельных этапов индивидуального развития сосудистых сплетений головного мозга (plexus choroids) миноги, рыб, земноводных, птиц, млекопитающих и человека позволило выявить этапы их морфогенеза и структурно-функционального становления гематоэнцефалического барьера.
Так, у хордовых основными закономерностями морфогенеза сосудистых сплетений головного мозга можно считать следующее: первичные сосудистые сплетения миноги являются простейшими образованиями и построены из эпителия. Последний представлен несколькими слоями. Сосуды лишь прилежат к нему.
У рыб начинают формироваться однослойные эпителиальные тяжи, окаймляющие примитивную соединительную ткань. Эпителиоциты резко не равноценны по высоте.
У амфибий ССГМ присутствуют во всех желудочках: боковых, третьем и четвертом. У них формируются не только первичные, но и вторичные ворсины. При этом сохранен план строения: однослойный эпителий расположен на базальной мембране. Клетки кубической формы, иногда уплощенные. В первых ядра округлые, во вторых – овальные. Все они содержат по 1–2 ядрышка. В соединительной ткани множественные сосуды микроциркуляторного русла. Наблюдаются единичные фибробласты.
У рептилий наибольшую функциональную нагрузку несут сосудистые сплетения боковых и четвертого желудочков. Именно они наиболее мощно представлены структурно. В этом классе хордовых эпителий представлен аналогично по сравнению с более низшими эволюционными формами. Но соединительнотканная прослойка содержит большее количество клеточных элементов, среди которых фибробласты, единичные фиброциты и пигментоциты.
У птиц закладка сосудистого сплетения формируется в боковых желудочках в виде многослойного эпителия с дальнейшей реорганизацией в однослойный. Порядок сходен с таковым у млекопитающих. Также наблюдается схожее строение в силу множественного ветвления ворсин в боковых желудочках головного мозга как у человека. Эпителий сосудистых сплетений однослойный кубический. Расположен на базальной мембране. Соединительнотканная прослойка представлена рыхлой соединительной тканью с сосудами микроциркуляторного русла и клеточными элементами.
Анализируя полученные морфометрические данные на примере ССГМ млекопитающих – мышей (рис. 1), можно отметить, что высота эпителиоцитов plexus choroids (хориоэпителиоцитов) 4-го желудочка во все сроки развития по размерам превышает размеры эпителиоцитов в боковом и 3-м желудочке (p < 0,05). Очевидно, это обусловлено функциональной необходимостью именно этого отдела центральной системы у млекопитающих. Plexus choroids боковых и третьего желудочков подобных изменений не обнаружено (p > 0,05).
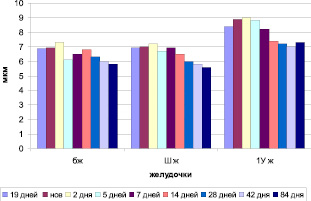
Рис. 1. Морфометрия сосудистых сплетений головного мозга мышей в онтогенезе (эпителий)
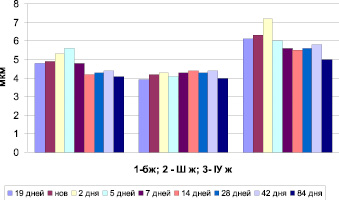
Рис. 2. Морфометрия сосудистых сплетений головного мозга мышей в онтогенезе (ядра)
У новорожденных мышей только в четвертом желудочке наблюдается увеличение высоты эпителия (p < 0,05). Эпителиоциты боковых и третьего желудочков практически не меняются.
По мере увеличения длительности постнатального периода до второго дня нарастает высота эпителиоцитов. Именно в этот период она достигает максимальных значений. Однако, к 5-ти дням в боковом желудочке наблюдается минимальная высота, в отличие от третьего и четвертого желудочков. Далее до 14 дня идет, вероятно, накопление секрета в эпителиоцитах боковых желудочках (p < 0,05). В третьем и четвертом желудочках, напротив, регистрируется снижение высоты эпителиальных клеток (p < 0,05). К 84 дню постнатальной жизни в боковых и третьем желудочках продолжается снижение высоты эпителиоцитов в отличие от эпителиоцитов четвертого желудочка (p < 0,05).
Другим объективным показателем морфофункциональной активности любой клетки является объем ядра. Так, на рис. 2 представлены результаты измерений объемов ядер в ходе онтогенеза. Очевидным является особое взаимоотношение между высотой эпителиоцита и размерами ядра в боковых желудочках на 5-й день постнатального развития, что, возможно, связано с активацией синтетической функции ядра в этот возрастной период. В то время как к моменту прозревания – 14 дней объем ядра значительно снижается, а высота эпителиоцита увеличивается.
Синтетическая активность ядер эпителия третьего желудочка колеблется в небольших пределах (p > 0,05), хотя эпителий имеет тенденцию к снижению высоты (p < 0,05).
В четвертом желудочке резкое увеличение синтетической активности ядер отмечается на 2-й день постнатальной жизни, которая уменьшается к моменту прозревания (14 день).
В постнатальном периоде у млекопитающих наименьшим изменениям подвергается объем ядер эпителиоцитов третьего желудочка, что свидетельствует об отсутствии необходимости интенсивного синтеза.
Нельзя не отметить разнонаправленные изменения объемов ядер бокового и четвертого желудочков. Так, если к 5-ти дням в боковом желудочке резко активируются синтетические процессы, то в четвертом, наоборот резко снижаются.
Ещё один скачок синтетической активности ядер сосудистых сплетений наблюдается в четвертом желудочке к 42 дням постнатальной жизни. Но к 84 дням во всех желудочках ядра снижают свою синтетическую активность.
Высота эпителия практически не меняется от новорожденного до 5 дней постнатального периода (p > 0,05).
Заключение
Исследование plexus choroids головного мозга позвоночных в сравнительном аспекте позволило установить ряд закономерностей, свидетельствующих об особенностях становления морфофункционального субстрата гематоэнцефалического барьера. Процессы дифференцировки наблюдаются во всех основных морфологических структурах, в частности в хориоэпителиоцитах.
Библиографическая ссылка
Шерышева Ю.В., Журавлева Г.Ф., Морозов В.Д. ФИЛОГЕНЕЗ СОСУДИСТЫХ СПЛЕТЕНИЙ ГОЛОВНОГО МОЗГА ПОЗВОНОЧНЫХ // Научное обозрение. Биологические науки. 2019. № 1. С. 67-71;URL: https://science-biology.ru/ru/article/view?id=1137 (дата обращения: 27.06.2026).