

 science-review.ru
science-review.ru
Естественное изреживание наиболее активно в возрасте 30–40 лет [1]. Принято считать, что слабые деревья отпадают и выживают самые крупные, которые используют освободившуюся территорию и увеличивают свои размеры за счет неё. Однако эти умозаключения, которые кажутся бесспорными, и разработанные на их основе Правила ухода за лесами [2] не подкреплены точными расчётами эффекта от их реализации, например в виде увеличения размеров остающихся деревьев спустя 10–20 лет после освобождения вблизи них территорий в результате отпада, гибели (или удаления) отставших в росте деревьев. Выяснение характера влияния дополнительной площади питания на увеличение размера оставшихся деревьев поможет прояснить проблему верификации теорий, основанных на якобы безусловной правильности так называемых «логических» моделей ухода за лесом, вытекающих и основанных на действии закона естественного изреживания лесных насаждений [3]. Если проверять эти модели в прямом эксперименте, то это будет убедительно; такие опытные рубки были, но результат получился совсем не тот, на который рассчитывали по крайней мере три поколения исследователей [4, 5]. Более того, расчеты конкурентного давления деревьев в молодняках сосны [6] и изучение площадей питания деревьев в спелых насаждениях показали, что на размер центрального дерева в микроценозе они влияют очень слабо – в пределах всего лишь от 0,2 до 11,0 % [6, 7].
Цель исследования – выяснить влияние площади питания дерева при её увеличении в результате отпада соседей в микроценозе на фоне естественных колебаний текущей густоты.
Материалы и методы исследования
Для изучения выбрали лесные культуры сосны в возрасте 55 лет 1Б класса бонитета с относительной полнотой 0,94, где заложили серию из 24 пробных площадей с нумерацией и определением диаметра деревьев через окружность ствола с точностью ± 0,1 см. В полевых условиях деревья наносили на абрис по прямоугольным засечкам с точностью ± 3–5 см и далее составляли карту их расположения в программе «ArcMap-ArcView». Для расчёта относительной полноты и запаса использовали «Стандартную таблицу полноты и запасов основных лесообразующих пород в Пермском крае» Г.С. Разина [7, с. 209]. Входом в неё служит так называемая «господствующая» высота [8], равная в среднем 28,4 м, и стандарт полноты для неё в наших таблицах [7] получается 45,7 м2/га, и стандарт запаса 578 м3/га. Полученные относительные полноты умножали на этот запас и получали запасы на 1 га на каждой пробе с округлением до десятков кубометров. Результаты приведены в табл. 1.
Таблица 1
Таксационные показатели культур сосны на пробных площадях (ПП)
|
№ п/п |
Площадь, га |
Живых деревьев, шт. |
Средние |
Полнота, м2/га |
Полнота относи-тельная |
Запас, м3/га |
||||
|
на ПП |
на 1 га |
Д, см |
Нср, м |
Нгосп, м |
на ПП |
стандартная |
||||
|
1 |
0,071 |
93 |
1310 |
20,8 |
27,5 |
28,4 |
39,9 |
45,7 |
0,87 |
500 |
|
2 |
0,071 |
106 |
1490 |
20,0 |
26,3 |
27,6 |
43,5 |
45,3 |
0,96 |
560 |
|
3 |
0,071 |
115 |
1620 |
19,0 |
26 |
27,3 |
44,0 |
45,2 |
0,97 |
560 |
|
4 |
0,071 |
109 |
1530 |
18,4 |
25,7 |
27,3 |
45,7 |
45,2 |
1,01 |
580 |
|
5 |
0,071 |
94 |
1320 |
18,8 |
25,8 |
27,3 |
42,3 |
45,2 |
0,94 |
540 |
|
6 |
0,071 |
90 |
1270 |
20,3 |
27,6 |
28,3 |
46,0 |
45,6 |
1,01 |
580 |
|
7 |
0,071 |
67 |
940 |
22,5 |
28 |
28,7 |
39,2 |
45,8 |
0,86 |
490 |
|
8 |
0,071 |
73 |
1030 |
23,1 |
28,1 |
28,7 |
46,5 |
45,8 |
1,02 |
590 |
|
9 |
0,071 |
79 |
1110 |
21,6 |
27,9 |
28,5 |
40,8 |
45,7 |
0,89 |
520 |
|
10 |
0,083 |
88 |
1060 |
21,5 |
27,8 |
28,2 |
38,4 |
45,6 |
0,84 |
490 |
|
11 |
0,087 |
98 |
1130 |
21,6 |
27,8 |
28,2 |
41,4 |
45,6 |
0,91 |
520 |
|
12 |
0,087 |
96 |
1100 |
22,0 |
27,9 |
28,3 |
42,1 |
45,6 |
0,92 |
530 |
|
13 |
0,081 |
98 |
1210 |
21,2 |
27,6 |
28,5 |
42,5 |
45,7 |
0,93 |
540 |
|
14 |
0,103 |
97 |
940 |
23,0 |
27,8 |
28,6 |
42,1 |
45,7 |
0,92 |
530 |
|
15 |
0,099 |
110 |
1110 |
22,0 |
27,7 |
28,5 |
42,1 |
45,7 |
0,92 |
530 |
|
16 |
0,093 |
110 |
1180 |
21,4 |
27,5 |
28,5 |
42,5 |
45,7 |
0,93 |
540 |
|
17 |
0,078 |
100 |
1400 |
20,2 |
27,3 |
28,3 |
44,9 |
45,6 |
0,98 |
570 |
|
18 |
0,0795 |
100 |
1260 |
20,7 |
27,4 |
28,5 |
43,3 |
45,7 |
0,95 |
550 |
|
19 |
0,073 |
109 |
1490 |
19,4 |
26,8 |
29 |
44,1 |
46 |
0,96 |
550 |
|
20 |
0,066 |
89 |
1350 |
20,4 |
27,3 |
29,1 |
44,4 |
46 |
0,97 |
560 |
|
21 |
0,075 |
93 |
1240 |
21,5 |
27,5 |
29,1 |
45,0 |
46 |
0,98 |
570 |
|
22 |
0,078 |
96 |
1230 |
20,6 |
27,4 |
29,1 |
41,1 |
46 |
0,89 |
520 |
|
23 |
0,079 |
99 |
1250 |
21,6 |
27,5 |
29,1 |
46,0 |
46 |
1,00 |
580 |
|
24 |
0,087 |
121 |
1390 |
20,5 |
27,5 |
29,1 |
46,1 |
46 |
1,00 |
580 |
|
Среднее |
0,079 |
97 |
1248 |
20,9 |
27,3 |
28,4 |
43,0 |
45,7 |
0,943 |
545 |
|
Колебания от среднего, % |
макс, |
130 |
110 |
103 |
102 |
108 |
101 |
108 |
108 |
|
|
мин, |
75 |
88 |
94 |
96 |
89 |
99 |
89 |
90 |
||
Для выстраивания полигонов питания выбрали 17 пробных площадей, имеющих высокую, среднюю и пониженную густоту. Число построенных полигонов питания вокруг живых деревьев составило 2219 шт., в том числе для возраста до 40 лет 1623 шт. и для возраста 41–55 лет 596 шт. полигонов. Возраст отпавших деревьев определяли по степени разложения пней и валежа, используемых для такой оценки их возраста [1, 8].
Результаты исследования и их обсуждение
Анализ результатов показал, что текущая густота культур от среднего значения 1248 шт/га колеблется в пределах 75–130 % с различием в 1,73 раза, и это открывает возможность разделения пробных площадей по этому фактору. Поэтому из 24 выбрали затем 17 пробных площадей с высокой, средней и пониженной густотой, где провели картирование живых (1623 шт.) и отпавших деревьев с расчетами их площадей питания для двух состояний: для возраста 30–40 лет, ориентируясь на следы отпавших деревьев, и для возраста 41–55 лет. Старые и новые площади питания с их перекрытием показаны на рисунке.
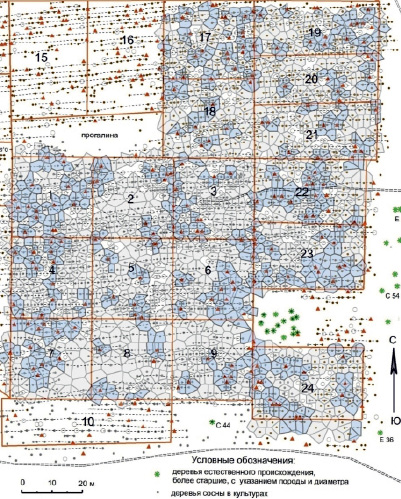
Старые (в возрасте 30–40 лет) и новые (в 41–55 лет) площади питания деревьев после естественного отпада деревьев на 17 пробных площадях в культурах сосны в возрасте 55 лет. Треугольные маркеры – старые пни и их остатки, овалы – сухостойные деревья
В работе использовали следующие обозначения площадей питания:
S0 – площади питания, которые не меня- лись;
S1 – старые площади питания в возрасте до 30–40 лет;
S2 – новые площади питания в возрасте 41–55 лет;
S2-20 – новые площади питания, увеличенные на 20 % и более;
Для расчетов провели сепарацию новых площадей питания S2 и выбрали только те, которые увеличились в сравнении с прежней площадью S1 на 20 % и более, что наблюдалось обычно при отпаде одного соседа. Их назвали «опытной» парной выборкой. В ней рассчитали средние значения S2-20, а также средние значения старых S1. Затем сформировали контрольную выборку; в неё подбирали деревья с неизменившейся площадью питания S0, но таким образом, чтобы среднее S0 в ней равнялось среднему значению S1 в опытной выборке.
После всех этих группировок можно было сравнивать средние диаметры стволов в контроле и в опыте. В теоретическом плане этот метод подбора данных для анализа сопоставим со слабыми рубками разреживания, но имеет преимущество в том, что не нужно ждать результатов с отсрочкой на 15–20 лет. В контрольные выборки подбирали деревья больше по их числу в густых местах в среднем на 44 % (в 1,25–1,53 раза), а в редких местах культур – на 27 % (в 1,06–1,43 раза). Результаты расчётов представлены в табл. 2.
Таблица 2
Статистические показатели площадей питания и диаметров стволов сосны у опытных и контрольных деревьев
|
№ п/п |
Густота шт/га |
Опытные деревья с увеличением площади питания на 20 % и более |
Диаметр ствола |
Отношение Д2 / Д0, % |
||||||||||
|
площадь питания в 30–40 лет (S1) |
площадь питания в 41–55 лет (S2) |
в опыте |
в контроле при средней S0 = S1 |
|||||||||||
|
N, шт. |
S1 , м2 |
± m |
СV, % |
S2 , м2 |
± m |
СV, % |
Д2 , см |
± m |
N, шт |
Д0 , см |
± m |
|||
|
Густые места |
||||||||||||||
|
3 |
1580 |
15 |
4,03 |
0,36 |
32 |
5,7 |
0,44 |
28 |
18,7 |
1,35 |
23 |
18,7 |
0,68 |
100,3 |
|
4 |
1530 |
17 |
4,05 |
0,28 |
26 |
5,9 |
0,40 |
26 |
19,8 |
1,17 |
26 |
18,1 |
0,65 |
109,7 |
|
2 |
1490 |
17 |
4,49 |
0,51 |
42 |
6,4 |
0,55 |
33 |
20,0 |
1,04 |
24 |
19,9 |
0,76 |
100,3 |
|
19 |
1490 |
19 |
4,55 |
0,47 |
42 |
6,7 |
0,55 |
34 |
20,5 |
1,18 |
28 |
18,4 |
0,62 |
111,3 |
|
17 |
1400 |
23 |
4,53 |
0,34 |
36 |
6,6 |
0,40 |
29 |
20,0 |
0,88 |
32 |
19,2 |
0,60 |
104,4 |
|
24 |
1390 |
20 |
5,46 |
0,42 |
33 |
7,9 |
0,57 |
30 |
19,8 |
0,86 |
25 |
20,26 |
0,8 |
97,7 |
|
20 |
1350 |
14 |
5,07 |
0,51 |
35 |
6,6 |
0,62 |
32 |
18,2 |
1,13 |
21 |
20,76 |
1,3 |
87,9 |
|
5 |
1320 |
13 |
4,06 |
0,28 |
23 |
5,8 |
0,42 |
24 |
18,4 |
1,03 |
20 |
18,5 |
0,83 |
99,9 |
|
1 |
1310 |
24 |
5,79 |
0,33 |
27 |
8,4 |
0,49 |
27 |
21,1 |
0,94 |
32 |
21,0 |
0,94 |
100,3 |
|
Число |
162 |
231 |
||||||||||||
|
Среднее |
18 |
4,67 |
32,8 |
6,67 |
29,3 |
19,61 |
19,42 |
101,0 |
||||||
|
Ср, взвеш, |
0,13 |
0,17 |
19,77 |
0,36 |
19,45 |
0,27 |
101,7 |
|||||||
|
Редкие места |
||||||||||||||
|
6 |
1270 |
23 |
5,3 |
0,7 |
46 |
7,5 |
1,1 |
42 |
21,7 |
1,1 |
28 |
21,6 |
0,9 |
100,5 |
|
18 |
1270 |
12 |
5,1 |
0,43 |
28 |
6,7 |
0,58 |
29 |
19,3 |
1,12 |
16 |
22,1 |
1,3 |
87,5 |
|
23 |
1250 |
25 |
6,1 |
0,38 |
31 |
8,5 |
0,56 |
33 |
22,3 |
0,89 |
33 |
19,9 |
0,74 |
111,8 |
|
21 |
1240 |
17 |
4,5 |
0,4 |
39 |
6,8 |
0,6 |
38 |
20,4 |
1,3 |
18 |
19,8 |
0,89 |
103,4 |
|
22 |
1230 |
23 |
5,0 |
0,3 |
29 |
7,1 |
0,4 |
26 |
18,3 |
0,7 |
30 |
19,6 |
0,7 |
93,4 |
|
9 |
1110 |
14 |
5,9 |
0,8 |
48 |
8,0 |
1,0 |
43 |
22,0 |
1,5 |
18 |
19,2 |
1,2 |
114,5 |
|
8 |
1030 |
7 |
6,3 |
0,8 |
33 |
8,9 |
1,0 |
27 |
20,4 |
1,6 |
10 |
23,2 |
0,9 |
88,1 |
|
7 |
940 |
17 |
5,84 |
0,4 |
29 |
8,84 |
0,7 |
32 |
22,1 |
0,9 |
20 |
21,7 |
1,2 |
101,9 |
|
Число |
138 |
173 |
||||||||||||
|
Среднее |
17 |
5,50 |
35,2 |
7,79 |
33,6 |
20,81 |
20,89 |
99,7 |
||||||
|
Ср, взвеш, |
0,20 |
0,27 |
20,89 |
0,41 |
20,64 |
0,35 |
101,2 |
|||||||
|
Число всего |
300 |
404 |
||||||||||||
|
В целом ср, взвеш, |
5,06 |
33,9 |
7,20 |
31,3 |
20,29 |
20,15 |
101,5 |
|||||||
Примечание. Ср. взвеш. – средневзвешенное значение.
В местах с высокой густотой на 9 пробных площадях сформировалась выборка из 162 деревьев, где площадь питания увеличилась от S1 = 4,67 до S2 = 6,67 м2. Увеличение составило в среднем 1,43 раза, с колебаниями по пробам от 1,3 до 1,46 раза. Заметим, что вариация площади питания при этом несколько снизилась: от 32,8 % у старых до 29,3 % у новых площадей питания. После получения дополнительной площади питания деревья в этой опытной выборке сформировали стволы со средневзвешенным диаметром 19,77 ± 0,36 см. В контроле средневзвешенный диаметр оказался 19,45 ± 0,27 см. Наблюдаемое превышение в 1,7 % недостоверно при t = 0,71 < < t0,95 = 1,98.
В местах с низкой текущей густотой на 8 пробных площадях сформировали выборку из 138 деревьев, где площадь питания увеличилась от S1 = 5,50 до S2 = 7,79 м2. Увеличение составило 1,42 раза, с колебаниями по пробам от 1,32 до 1,49 раза. Вариация площадей питания также снизилась незначительно: от 35,2 % у старых и до 33,6 % у новых площадей. Деревья к 55 годам сформировали стволы со средневзвешенным диаметром 19,89 ± 0,41 см в опыте и 20,64 ± 0,35 см в контроле. Превышение составило 1,2 % и также было недостоверно, причём если суммировать просто средние диаметры по пробам, то превышение вообще исчезает (табл. 2).
В целом во всей опытной выборке из 300 деревьев размер площади питания после естественного отпада деревьев увеличился от 5,06 до 7,2 м2, или в 1.42 раза. В контрольной выборке из 404 деревьев отпада не было и площадь питания осталась равна в среднем 5,06 м2. В сравнении с контролем спустя примерно 15–20 лет средний диаметр центрального дерева в «опытных» микроценозах увеличился лишь на 1,5 %. Несмотря на большие выборки, это слабое превышение оказалось в итоге недостоверным.
Полученные результаты слишком серьёзны, чтобы оставить их без детального обсуждения. Во-первых, уже давно известно, что решающим фактором для успешного развития деревьев является не «регуляция густоты в период дифференциации», как это пишется в учебниках, а густота начальная, т.е. густота «на старте» в возрасте около 10 лет. В это время начинают функционировать ранговый закон роста деревьев в молодняках Е.Л. Маслакова (1984) [9] и одновременно с ним – закон развития одноярусных древостоев в зависимости от начальной густоты Г.С. Разина [3]. Фактор густоты на уровне насаждения действует на порядок сильнее, чем на индивидуальном уровне, где обычно используют обратную густоте величину – площадь питания. Затем вступает в действие «Закон генетического гомеостаза частот правых и левых форм деревьев» А.М. Голикова [10], по которому при достижении высокой полноты в насаждении начинают лидировать толерантные к конкуренции деревья правой формы, и они в малой степени нуждаются в дополнительной площади питания. При этом отмирают деревья левой формы, плохо переносящие высокую густоту [7, 10].
Неудивительно, что если не учитывать действие законов Маслакова, Разина и Голикова, первые два из которых известны уже более 30 лет, но до сих пор почему-то не используются, то применяемые для возраста 30–55 лет «логические» модели ухода за лесом, имманентно содержащиеся в Правилах ухода за лесами [2], разработанных на основе только одного закона естественного изреживания, воздействуют лишь на часть процессов и работают как квазимодели. Поэтому их практические воплощения скомпрометировали и их идею, и сами Правила ухода за лесами. Эти устаревшие модели должны быть преодолены как не отвечающие современному развитию лесных наук. Они существуют в некотором смысле уже как фантомы теорий [11], но у сторонников коммерческих рубок ухода [12] постоянно возникают попытки как-то подтвердить их легитимность, используя некие новые идеи внутри старой идеологемы «борьбы видов за существование» Ч. Дарвина, упрощённо объяснявшей процесс естественного изреживания лесных насаждений [7].
Поэтому, если знать о том, что в насаждениях с высокой полнотой в возрасте 30–40 лет всегда начинают преобладать правые формы деревьев, толерантные к высокой конкуренции, то становится понятной очень слабая реакция таких деревьев на увеличение площади их питания к 55 годам, так как даже «редкие» места в изученном массиве культур имеют полноту выше 0,8. Ныне исследователи признают, что известных факторов уже недостаточно для математических описаний и моделирования пространственно-временной структуры древесных сообществ [13, 14]. В связи с этим настоятельно необходим поиск иных абиотических факторов, влияющих на развитие деревьев в микроценозах. Возможно, они будут связаны с действием пока малоизученных глубинных энергий Земли.
Заключение
1. Установлено, что в совокупной выборке из 300 деревьев с увеличением площади питания в 1,42 раза после естественного отпада деревьев вблизи них в возрасте примерно 30–40 лет диаметр ствола к возрасту 55 лет увеличился недостоверно и лишь на 1,5 % в сравнении с контролем (404 дерева), где площади питания остались прежними.
2. Разделение совокупной выборки с увеличением площади питания центрального дерева на совокупность в густых (162 дерева) и в редких местах культур (138 деревьев) не выявило достоверных различий между ними по превышению диаметра ствола у дерева в центре полигона питания в сравнении с контрольными деревьями.
3. В связи с действием закона генетического гомеостаза частот правых и левых форм деревьев А.М. Голикова [10] и в процессе достижения насаждением высокой полноты в нём начинают преобладать толерантные к конкуренции деревья правой формы, и они в малой степени нуждаются в дополнительной площади питания. Поэтому естественное изреживание культур и получение деревьями дополнительной площади питания в период от 30 до 40 лет эта площадь, как результат изреживания, в течение 15–25 лет почти не повлияла на рост по диаметру оставшихся деревьев. В связи с этим интенсивные рубки разреживания в возрасте старше 30 лет практически бесполезны и должны быть заменены на модели с активной регуляцией густоты в возрасте 10–20 лет.
Библиографическая ссылка
Рогозин М.В. ВЛИЯЕТ ЛИ ЕСТЕСТВЕННОЕ ИЗРЕЖИВАНИЕ В КУЛЬТУРАХ СОСНЫ НА РАЗМЕР ДЕРЕВЬЕВ? // Научное обозрение. Биологические науки. 2021. № 4. С. 44-49;URL: https://science-biology.ru/ru/article/view?id=1243 (дата обращения: 02.08.2026).
DOI: https://doi.org/10.17513/srbs.1243