

 science-review.ru
science-review.ru
В современных исследованиях лесоводы все более склоняются к тому, что размеры деревьев зависят в большей степени не от наследственности и внутривидовой конкуренции, а от экологической неоднородности биотопа [1–3]. Так в 184-летнем насаждении сосны обыкновенной 1 класса бонитета нами изучались 18 кандидатов в плюсовые деревья (далее кандидаты). Средняя площадь их питания была больше, чем у обычных деревьев (на 19,7 %), однако отличие было недостоверно. Возраст определяли по кернам у 40 деревьев, и у кандидатов он колебался от 174 до 193 лет. Вероятно, выдающиеся размеры у молодых кандидатов были обусловлены либо генетически, либо тем, что имеются особо благоприятные места, где деревья поселились позднее и не только догнали соседей, но перегнали их и достигли выдающихся размеров. Высказано предположение, что такие места могут быть связаны с влиянием малоизученных энергий Земли [3].
Изучение таких мест под названием «геобиологические активные зоны» началось в 1974 г. [2]. Согласно этим исследованиям прогалины в древостое приурочены к дискомфортным зонам, а скопления деревьев – к зонам, благоприятным для роста. В результате И.С. Марченко впервые поколебал теорию лесоводства, основанного на принципе равномерного размещения деревьев. В 1998 г. в учебнике «Нетрадиционное лесоводство» он предложил заменить его на принцип группового размещения деревьев [4].
Начиная с 2008 г. изучение таких зон началось в Пермском крае М.В. Рогозиным [3, 5]. Он выделил дискретные зоны с благоприятным влиянием на растения, с размерами 1, 3, 8, 16, 32 и 55. Определялся их размер и точное положение, а также выстраивались их сети, и они получили новое название – «малые геоактивные зоны» (далее МГА-зоны или просто зоны). Так, в упомянутом выше 184-летнем сосняке [3] «младшие» зоны размером 1 и 3 м встречались с частотой 220–250 шт/га; сосна занимала их на 90 % и сохранялась в 39 раз лучше, повышая объём ствола на 46 %. Далее, после анализа расположения МГА-зон вблизи крупных деревьев других пород, был сделан вывод, что крупные деревья являются их объективными фитоиндикаторами. Сети и сами зоны с размерами 1 и 3 м занимают 38 % территории, и их можно использовать для выращивания леса как энергетический каркас, где деревья подпитываются глубинными энергиями Земли. В частности, холодной и затяжной весной эти зоны увеличили грунтовую всхожесть семян ели в 7 раз (!), и поэтому они могут быть использованы для выращивания посадочного материала в условиях меняющегося климата. По-видимому, лесные породы эволюционировали с использованием их энергий; во всяком случае, все изученные виды деревьев увеличивали на них свое долголетие, размеры, семеношение и даже меняли наследуемость с положительной на отрицательную [3, с. 17]. Зоны неоднородны и состоят из поясов ингибирования, депрессии и комфорта [3, с. 174].
Начиная с 2016 г. началось использование морфоструктурного анализа космоснимков, геокарт и другой информации с ранжированием плотности так называемых линеаментов, отражающих трещиноватость погребенного фундамента. В Пермском крае к 2012 г. их было выделено 49 тыс. [6]. По степени их плотности выделено более 100 геодинамических активных зон с чрезвычайно высокой активностью (6-й ранг плотности линеаментов), которые трассировали тектонические разломы; наряду с ними имеются и спокойные места (1–2-й ранг плотности линеаментов). Эти зоны проявляют себя через геохимические, магнитные и гравитационные аномалии, химический состав воды родников и т.д. Они начинаются с размера 0,7 км и достигают десятков километров. В целом территория края представляет собой сеть из пресекающихся разломов разного уровня [6].
Учёт плотности линеаментов впервые позволил обнаружить связь этого фактора с увеличением плеч миграции МГА-зон [3, с. 9]. Затем, рассматривая их на фоне новых открытий в космогеологии [7], было установлено, что сети из МГА-зон с размерами 1 и 8 м в горах были ориентированы в одном направлении, а на равнине их ориентация была рассогласована на 30°. На ряде примеров было показано, что это вызывалось наличием разломов разного генезиса: линейных, кольцевых и спиральных, для нахождения которых используют специальный геоструктурометрический анализ [7, 8]. При таком анализе проводят математическую обработку космоснимков с определением параметров для каждого из миллионов пикселей в разных спектральных каналах на основе научного открытия российского учёного Ю.И. Фивенского [9].
Изучаемые нами МГА-зоны встречаются повсеместно и имеют постоянные размеры, в отличие от кольцевых структур Ю.И. Фивенского [10]; по-видимому, это отражает их одинаковую мощность и частотные характеристики. Влияние таких зон на почву и её биоту неизвестно. Физическая природа МГА-зон представляет собой настоящую загадку, и разгадать её можно только чувствительными приборами. В настоящее время высказывается ряд гипотез о природе энергий Земли, формирующих геологический ландшафт [11, 12], и мы наблюдаем их наиболее выразительно, как разрушительную силу. Но тогда должны быть и силы созидающие, т.е. те силы, которые сохраняют горы, и «молчание» горных вершин – самое интригующее. Для сильных энергий есть карты гравитационных полей, аномалий магнитного поля, сейсмоактивности и другие [6, 7, 10]. Однако действие слабых сил почти не изучено, и часть их можно отнести, по-видимому, к тонким энергиям.
О тонких энергиях применительно к Человеку в интернете масса публикаций. Но и наша Земля – «энергетически живой» мегаорганизм, и энергий у неё множество, поэтому отказать ей в наличии тонких энергий было бы несправедливо. Их измерение, однако, наталкивается на сильные помехи. Типичная помеха – сигнал геомагнитного поля Земли в датчиках магнитометра [11]. Однако вполне может быть, что даже самые совершенные магнитометры пока не смогут улавливать сигналы этих тонких энергий, частоты которых очень высокие.
Не углубляясь в их физические особенности, отметим, что растения «знают» и используют эти тонкие энергии. В точке, где растёт дерево, одновременно действует множество излучений, поэтому А.М. Гореловым с соавт. было предложено называть его «геогенным» и измерять суммарно биолокацией [12, c. 98]. По-видимому, их следует искать в диапазоне излучений с колебаниями 70–200 КГц, а также в диапазоне 680–900 ТГц, которые будут резонировать с биополями таких деревьев как береза, дубы, калина, клены, липа, метасеквойя, ольха чёрная, осина, сосна обыкновенная, тополя, ясень [12, с. 70].
Новое направление исследований с изучением среды обитания растений эниологией [12] с поддержкой дистанционными методами зондирования Земли [7, 8, 10] позволит продвинуть наши знания о лесах на новый уровень, где геофитоценология и лесная биофизика [13] займут, наконец, своё достойное место.
Таковы краткие итоги исследований геоактивных зон на стыке геологии и лесоведения. Они позволят здесь лучше понять результаты наших исследований о локализации плюсовых деревьев, которые будут связаны с пространственным размещением геоактивных зон.
Цель исследования – выяснить, в какой мере малые геоактивные зоны обуславливают формирование плюсовых деревьев в культурах сосны к возрасту 55 лет.
Материалы и методы исследования
Исследования проводились в 55-летних культурах сосны 1Б класса бонитета полнотой 0,94 на супесчаной почве в кв. 43 Нижне-Курьинского участкового лесничества Пермского городского лесничества. В них ограничили визирами однородный участок размером 80×80 м и пронумеровали все живые деревья и нанесли их на план с точностью ± 5–10 см. План оцифровали в программе «ArcMap-ArcView», после чего участок виртуально разделили на 9 пробных площадей, где вокруг деревьев выстроили полигоны их питания. Затем в полевых условиях с привязкой к центрам оснований стволов на план нанесли малые геоактивные зоны трёх типов с размерами 1, 3 и 8 м. Методика выстраивания полигонов питания и биолокационный метод нахождения центров геоактивных зон были описаны ранее [3].
Результаты исследования и их обсуждение
После перечислительной таксации и первичного анализа результатов [14] в культурах были выделены плюсовые деревья (66 шт.) с превышением диаметра на 30 % и более. Часть из них имела раздвоение ствола, толстые сучья и другие недостатки, но нам был важен только их крупный размер как показатель виталитета. Поэтому использование термина «плюсовые» здесь условное, и под ним понимается только наличие у дерева указанного превышения диаметра без других качеств, присущих плюсовым деревьям.
В предыдущей работе [14] различие между средней площадью питания у 66 плюсовых (8,27±0,56 м2) и у прочих деревьев (6,11±0,11 м2) было достоверно при t = 3,8 > t0.01 = 2,6. Из них 24 шт. (36 %) сформировались на меньшей, и остальные – на большей площади питания, чем её среднее значение, равное 6,28 м2 для всех 823 деревьев на этой территории. В данной работе мы рассмотрим только эти 24 плюсовых дерева с малой площадью питания. Для них можно утверждать, что они сформировались крупными либо благодаря их генетическим задаткам, либо на них повлияли какие-то иные факторы, улучшающие их рост. На участке они встречаются достаточно равномерно (рис. 1).
Далее рассмотрим влияние трёх типов геоактивных зон с размерами 1, 3 и 8 м, так как именно они были наиболее благоприятны для роста большинства лесных пород. Вставим слои карт с этими зонами на предыдущий слой карты с плюсовыми деревьями (рис. 2).
Анализ рисунка показывает, что 24 плюсовых дерева растут в следующих местах:
− 4 шт., или 17 %, растут на свободных от зон и полос-цепей местах;
− 6 шт., или 25 %, находятся внутри полос-цепей, образуемых зонами размером 1 м;
− 14 шт., или 58 %, растут внутри или на внешнем радиусе зон размером 1, 3 и 8 м.
Получается, что среди растущих на малой площади питания плюсовых деревьев лишь 17 % не имели подпитки энергиями Земли и формировались за счёт собственных усилий при отсутствии энергетической составляющей их среды обитания. Впрочем, нельзя исключить, что на них воздействовали ещё и другие, «старшие» зоны размером 16, 32 и 55 м, которые здесь пока не учитывались; их биолокацию мы проведём в следующем полевом сезоне. Эти старшие зоны были в целом нейтральны для сосны в возрасте 184 года, хотя на зонах размером 16 м она всё же сформировала 4,6 % плюсовых деревьев [3, с. 124]. В связи с этим они будут весьма интересны для генетики как организмы, «сделавшие себя сами» без подпитки энергиями зон размером 1 и 3 м, на которых формируется большинство крупных деревьев самых разных пород. По отношению ко всем плюсовым деревьям (66 шт.) их частота здесь 6 %, а к общему числу 823 живых деревьев их доля равна 0,5 %.
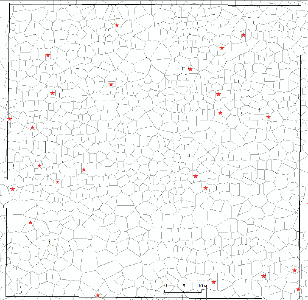
Рис. 1. Полигоны питания в культурах сосны на территории 80×80 м и 24 плюсовых дерева, имеющих малые площади питания
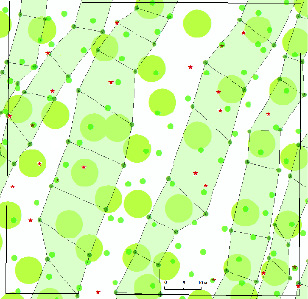
Рис. 2. Плюсовые деревья с малой площадью питания, благоприятные зоны и полосы-цепи на территории 80×80 м
Если же не учитывать цепи-полосы, энергия которых, как мы полагаем, менее интенсивна и значимо влияет на рост деревьев лишь в молодости [3, с. 105–113], то тогда без подпитки энергией МГА-зон остаётся 10 плюсовых деревьев. От общего числа деревьев это составит 1,2 %. Они также представляют интерес для лесной селекции и генетики как деревья, которые используют менее интенсивные энергии Земли.
Обсуждая результаты, следует сравнить их с анализом роста деревьев в 184-летнем сосняке, где из 44 кандидатов в плюсовые деревья 89 % сформировались на указанных выше трёх типах МГА-зон. Это позволяет сделать прогноз, что из наших 66 плюсовых деревьев через 100–120 лет останется только та часть, которая растёт на зонах указанных типов.
Таким образом, фактор местообитания плюсовых деревьев надо рассматривать намного шире, чем это принято, так как некоторые места биотопа связаны с мощным влиянием энергий Земли, выходящих через малые геоактивные зоны. Этот энергетический фактор, по-видимому, влияет и на почву, химические процессы в ней и активность микроорганизмов. Для его изучения необходимы новые технологии, которые позволят отделять абиотические факторы и точнее рассчитывать долю влияния генотипа. Тогда лесная биогеофизика по праву займёт своё достойное место среди лесных наук [13], а поселение деревьев на скалах и их успешный рост почти без почвы [15] получит, наконец, внятное объяснение.
Заключение
1. В культурах сосны в возрасте 55 лет на малых площадях питания 58 % плюсовых деревьев сформировались при подпитке энергиями геоактивных зон размером 1 и 3 м.
2. Внутри образуемых зонами размером 1 м пространства полос-цепей, с выходом менее интенсивных энергий Земли, образовалось 25 % плюсовых деревьев, имеющих малую площадь питания.
3. На свободных от полос-цепей и зон размером 1, 3 и 8 м местах растёт 17 % плюсовых деревьев с малой площадью питания. Они могут быть интересны для генетики как деревья, рост которых не зависел от конкурентного давления соседей и подпитки энергиями Земли, наиболее благоприятными для большинства видов деревьев.
4. При учёте влияния энергий, действующих через геоактивные зоны размером от 1 до 8 м, можно выделить уникальные плюсовые деревья (0,5–1,2 % от общего числа деревьев), которые выросли на малой площади питания без подпитки теми типами энергий, на которых формируется большинство крупных деревьев. Возможно, такие генотипы нужны для выживания вида в случае глобальных катаклизмов, например при смене полюсов Земли.
Автор благодарит за разрешение на исследования в Пермском городском лесничестве директора Галанову Антонину Александровну и лесничего Васильевых Геннадия Павловича.
Библиографическая ссылка
Рогозин М.В. ПЛЮСОВЫЕ ДЕРЕВЬЯ СОСНЫ И ГЕОАКТИВНЫЕ ЗОНЫ // Научное обозрение. Биологические науки. 2022. № 1. С. 5-9;URL: https://science-biology.ru/ru/article/view?id=1251 (дата обращения: 16.06.2026).