

 science-review.ru
science-review.ru
Энтомофауна Узбекистана очень разнообразна и включает много эндемичных для Центральной Азии видов. Одними из таких видов являются представители рода Machozetus Chaudoir, 1850 из семейства жужелиц (Carabidae). Этот род жужелиц является эндемическим родом Центральной Азии и включает всего два вида: Machozetus lehmanni Menetries, 1848 и Machozetus concinnus C.A. Dohrn, 1885. Они отмечены в пустынных и предгорных районах Узбекистана, Туркменистана, Таджикистана и Северного Афганистана [1]. В последние годы на основе музейных материалов было доказано распространение этих видов в Иране [2]. Виды рода Machozetus являются специализированными фитофагами, питающимися семенами растений, в основном ферулы [3]. Machozetus lehmanni, как своеобразный монофаг зонтичных растений, отмечен в Заповеднике Репитак (Туркменистан) на песчаных барханах [4].
Жизнь и развитие Machozetus lehmanni проходит в специальной норе в почве. Самки роют норы в вертикальном направлении на глубине до 4 м, где откладывают по одному яйцу и запасают семена ферулы для кормления личинок [3]. Личинки развиваются в почве и впервые были описаны И.Х. Шаровой и К.В. Макаровым. Личинки 3 возраста до 34 мм, тело покрыто светлыми и мелкими волосками, глаза неразвиты. Эти особенности свидетельствует об их малоподвижном и скрытом образе жизни [5]. А.Ш. Хамраевым (2013) в Харезмской области и Республике Каракалпакистан Machozetus lehmanni и Machozetus concinnus отмечаются как представители подсемейства Nebrinae [6].
Хотя в вышеперечисленных исследованиях имеются сведения о распространении видов рода Machozetus, их морфологические и биологические особенности остаются особо не изученными. У насекомых морфологические особенности, обеспечивающие способность к расселению, эффективность поиска пищи и питания, определяют их взаимодействие с окружающей средой [7, 8]. В последние годы широкое распространение получил подход оценки структуры популяций насекомых по морфометрическим признакам [9, 10]. На примере агроценозов и урбанизированных ландшафтов выявлены значительные вариации морфометрических показателей у жужелиц. Например, C. granulatus уменьшается в размерах при обитании в пригороде, а C. cancellatus – в городе [11]. И даже фенотипическая пластичность самок и самцов на действие какого-либо фактора среды у жужелиц одного и того же вида может быть различной [12].
Хотя некоторые морфологические особенности Machozetus lehmanni были сообщены нами ранее [13], в этой статье приводятся данные по морфологии и морфометрическим показателям, а также вариациям (в том числе на разных участках ареала) данного вида.
Необходимо отметить, что в последнее время происходит значительное сокращение естественных местообитаний одного из ценных лекарственных растений – ферулы вонючей. Так как виды рода Machozetus являются специализированными фитофагами растений ферулы, детальное изучение как их морфо-биологических, так и популяционно-экологических особенностей, может иметь важное хозяйственное значение.
Материалы и методы исследования
Сбор материала проводился в пустынных регионах Бухарской, Кашкадарьинской и Навоийской областей Узбекистана в 2020–2021 гг. (рис. 1). Сбор материала проводился весной и летом методом ручного сбора, в основном ночью, так как жуки наиболее активны в ночное время. Собранных жуков фиксировали этилацетатом и раскладывали в ватные матрасики. Дальнейшее изучение морфологии жуков проводили в энтомологической лаборатории биологического факультета СамГУ. С материалом можно ознакомиться в энтомологической коллекции университета, где хранится весь собранный материал.
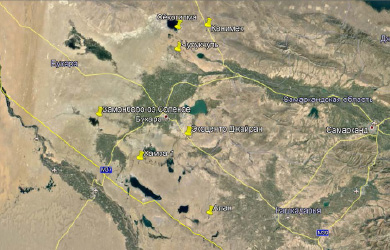
Рис. 1. Точки сбора материала
Изучение морфометрических показателей проводили с помощью мерного бинокулярного микроскопа МВС-9 с мерной линейкой.
Исследованы следующие морфометрические показатели: ДГ – длина головы, ШГ – ширина головы, РГ – расстояние между глаз, ДП – длина переднеспинки, ШП – ширина переднеспинки, ДЭ – длина элитры, ШЭ – ширина элитры и ДТ – общая длина тела (ДГ+ДП+ДЭ) (рис. 2). Дополнительно изучена пропорции разных частей тела: ДГ/ДТ, ШГ/ДТ, РГ/ДТ, ДП/ДТ, ШП/ДТ, ДЭ/ДТ, ШЭ/ДТ, ШП/ДП и ШЭ/ДЭ.

Рис. 2. Схема морфометрических измерений: 1–2 – длина головы – ДГ, 2–3 – длина переднеспинки – ДП, 3–4 – длина элитры – ДЭ, 5–6 – ширина элитры – ШЭ, 7–8 – ширина переднеспинки – ШП, 9–10 – ширина головы – ШГ, 11–12 – расстояние между глаз – РГ
Статистическая обработка полученных данных проводилась со стандартным пакетом Statistica 10. При этом были рассчитаны среднеарифметические значения, стандартное отклонение, стандартная ошибка, коэффициент вариации и коэффициент корреляции между размерами разных частей тела.
Результаты исследования и их обсуждение
Виды рода Machozetus Machozetus lehmanni и Machozetus concinnus являются морфологически очень похожими видами, и их разница в основном проявляется в размерах тела. В исследованных территориях численность популяции этих двух видов существенно различается. Если Machozetus lehmanni было зарегистрировано во всех семи исследованных точках, то Machozetus concinnus выявлено только в двух точках (Экоцентр «Жайран», Бухарская обл., и пустыня Алан района Минишкор Кашкадарьинской обл.) и малой численностью.
Machozetus concinnus – жук блестяще-черного цвета, длиной тела 20,2–25,5 мм. По морфологическим признакам очень похож на Machozetus lehmanni (рис. 3). Так как численность этого вида невысокая (всего собрано около 20 экземпляров) и собранный материал не подлежит статистической обработке, ниже приводим морфологическое описание только Machozetus lehmanni.

Рис. 3. Виды рода Machozetus Chaudoir, 1850: А – Machozetus lehmanni; Б – Machozetus concinnus (ориг.)
Machozetus lehmanni – крупный жук блестяще-черного цвета. Длина тела 29,6–42,8 мм. Голова крупная, вместе с челюстями длина головы составляет 10,7–17,5 мм, а ширина вместе с глазами – 7–11,3 мм, с очень сильно развитыми челюстями. Глаза крупные, черные или белые, в большинстве случаев черные глаза с белыми пятнами разных размеров в центре. Эти белые пятна иногда в центре глаз, иногда очень расширенные, в других случаях полностью охватывают глаза. В последнем случае глаза бывают белыми. Расстояние между глазами, в зависимости от ширины головы, составляет 6,3–9,6 мм. Усики 11-члениковые, 1-й членик очень крупный, 2-й членик в 1,5 раза короче остальных последующих члеников. 1–2-е членики усиков блестящие и имеют щетинки, начиная с 3-го членика, все остальные покрыты мелкими рыжими волосками. На кончике каждого членика расположены несколько щетинок – сенсилл.
Ротовой аппарат сильно развит. Верхняя губа покрыта рыжими волосками. Верхние челюсти крупные. Нижнечелюстные щупальца трёхчленные, черные, блестящие, кончик каждого членика желтоватый. 1-й членик крупный и толстый, в 1,5 раза длиннее 2-го членика. 3-й членик короткий, расширенный от основания к кончику, плоский. Щупальца нижних губ также трёхчленные, основной членик короткий, 2-й членик в 4–5 раза длиннее основного, 3-й членик в 1,5 раза длиннее основного, расширенный и плоский, кончик красновато-коричневый. Сама нижняя губа блестящего желтовато-коричневого цвета.
Ноги сильно развиты, бегательные, длинные, блестяще-черного цвета, покрыты коричневыми щетинками. Щетинки образуют плотные ряды по бокам голени.
Переднеспинка своеобразной сердцевидной формы, но нижняя часть не заострённая, всегда шире головы, в центре по вертикали проходит борозда. Длина переднеспинки – 5–7,1 мм, а ширина – 12–15,7 мм.
Элитра, как у большинства крупных жуков, сильно хитинизированная и сросшаяся. Блестяще-черного цвета. Места соединения двух надкрылий в виде узкой борозды. На каждой элитре имеются по 7–8 узких бороздок, которые делят надкрылия на полосы. Центральная бороздка, которая расположена между двумя надкрыльями, немного шире остальных. Ширина элитры составляет 12–15,7 мм, а длина – от 13,3 до 20,2 мм.
Вариация исследованных морфометрических показателей внутри популяции не очень высокая и является стабильными признаками, так как для нестабильных признаков коэффициент вариации должен быть более 33,3 %. Статистический анализ данных показал, что наиболее вариабельной является длина головы (Cv = 9,22), а наименьшее значение коэффициента вариации (Cv = 5,51) наблюдается для ширины надкрылий (табл. 1).
Как известно, размеры разных органов тесно взаимосвязаны. Однако степени зависимости в размерах различных органов могут быть неодинаковыми. Для выяснения степени зависимости провели корреляционный анализ между морфометрическими показателями измеренных частей тела жуков (табл. 2).
Высокая корреляционная зависимость выявлена между шириной головы (ШГ) и расстоянием между глаз (РГ), между шириной переднеспинки (ШП) и шириной надкрылий (ШЭ), между длиной надкрылий и общей длиной тела (r = 0,9). Наименьшая коррелятивная зависимость выявлена между длиной головы (ДГ) и длиной переднеспинки (ДП) (r = 0,31). Также слабая зависимость отмечена между длиной переднеспинки (ДП) и шириной головы (ШГ), между длиной переднеспинки (ДП) и расстоянием между глаз (РГ) (r = 0,50). В общем, длина переднеспинки проявляет слабую зависимость от морфометрических показателей головной части.
Таблица 1
Морфометрические показатели жука Machozetus lehmanni и их вариации (n = 100)
|
Части тела |
Мах |
Мin |
Средне-арифметическое, M |
Стандартное отклонение, sd |
Стандартная ошибка, m |
Коэффициент вариации, Cv, % |
|
ШГ |
11,3 |
7 |
9,5 |
0,77 |
0,08 |
8,10 % |
|
ДГ |
17,5 |
10,7 |
13,7 |
1,26 |
0,14 |
9,22 % |
|
РГ |
9,6 |
6,3 |
7,6 |
0,62 |
0,07 |
8,15 % |
|
ШП |
13,3 |
10 |
11,7 |
0,69 |
0,07 |
5,95 % |
|
ДП |
7,1 |
5 |
6,0 |
0,39 |
0,04 |
6,57 % |
|
ШЭ |
15,7 |
12 |
13,6 |
0,75 |
0,08 |
5,51 % |
|
ДЭ |
20,2 |
13,3 |
17,3 |
1,17 |
0,13 |
6,77 % |
|
ДТ |
42,8 |
29,6 |
36,9 |
2,38 |
0,26 |
6,46 % |
|
ШГ/ДТ |
0,29 |
0,23 |
0,26 |
0,01 |
0,001 |
4,94 % |
|
ДГ/ДТ |
0,41 |
0,32 |
0,37 |
0,02 |
0,002 |
4,92 % |
|
ШП/ДТ |
0,36 |
0,3 |
0,32 |
0,01 |
0,001 |
3,72 % |
|
ДП/ДТ |
0,18 |
0,14 |
0,16 |
0,01 |
0,001 |
5,35 % |
|
ШЭ/ДТ |
0,43 |
0,34 |
0,37 |
0,01 |
0,001 |
3,60 % |
|
ДЭ/ДТ |
0,51 |
0,43 |
0,47 |
0,01 |
0,002 |
2,97 % |
|
РГ/ДТ |
0,24 |
0,19 |
0,21 |
0,01 |
0,001 |
5,55 % |
|
ШП/ДП |
2,32 |
1,73 |
1,96 |
0,10 |
0,01 |
5,11 % |
|
ШЭ/ДЭ |
0,98 |
0,71 |
0,79 |
0,04 |
0,004 |
4,85 % |
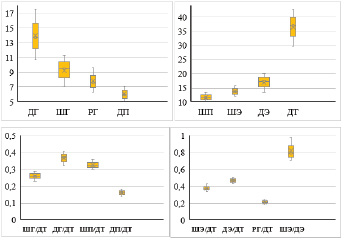
Рис. 4. Сдвиг в значениях морфометрических показателей у Machozetus lehmani (ед. изм. – мм,  – среднеарифметическое значение, – станд. ошибка, Т – станд. отклонение)
– среднеарифметическое значение, – станд. ошибка, Т – станд. отклонение)
Таблица 2
Корреляционная зависимость (r) между размерами разных частей тела у Machozetus lehmani (n = 100)
|
Части тела |
ШГ |
ДГ |
РГ |
ШП |
ДП |
ШЭ |
ДЭ |
ДТ |
|
ШГ |
||||||||
|
ДГ |
0,79 |
|||||||
|
РГ |
0,90 |
0,70 |
||||||
|
ШП |
0,85 |
0,70 |
0,84 |
|||||
|
ДП |
0,50 |
0,31 |
0,50 |
0,69 |
||||
|
ШЭ |
0,77 |
0,69 |
0,76 |
0,90 |
0,66 |
|||
|
ДЭ |
0,61 |
0,56 |
0,57 |
0,71 |
0,69 |
0,74 |
||
|
ДТ |
0,80 |
0,85 |
0,73 |
0,83 |
0,67 |
0,84 |
0,90 |
Хотя все исследованные жуки были собраны в схожих пустынных биотопах, провели сравнительный анализ морфометрических показателей жуков из наиболее отдалённых участков. Для этого была выбрана пустыня Алан (Кашкадарьинская область, 38048’26.69»С, 64059’06.65» В) и Чурукчул (Бухарская область, 40025’29.43»С, 640 42’55.88»В), расстояние между которыми составляло около 180 км. Из каждого участка измеряли по 30 случайных жуков. Полученные результаты показали некоторые различия в морфометрических показателях жуков (рис. 5).
Более крупными оказались жуки из участка Чурукчул, длина тела которых составляла 42,8 мм (±2,31), а у жуков из участка Алан – 39,1 мм (±2,3).
Интересен тот факт, что разница в размере тела в основном обеспечивается более крупными размерами головы. Так, среди измеренных морфометрических показателей существенные различия наблюдались только по размерам головы, а остальные показатели были практически на одном уровне. Вероятной причиной такой тенденции могут быть свойства (плотность) песка в исследованных участках, так как жуки Machozetus lehmani роют глубокие ходы в почве, там откладывают яйца и ухаживают за личинками, или же особенности кормового растения (соцветий и семян ферулы).
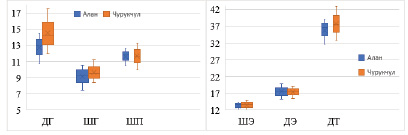
Рис. 5. Сравнение морфометрических показателей Machozetus lehmani, выявленных на двух наиболее отдалённых участках исследований (ед. изм. – мм, n = 30)
Хотя в некоторых работах для других видов жужелиц размеры тела различались в зависимости от растительного покрова [14]. Однако это обстоятельство требует дальнейшего детального изучения.
Заключение
Таким образом, Machozetus lehmanni и Machozetus concinnus очень близкие по морфологическим особенностям виды, но сильно различаются по размеру тела. В настоящей работе проанализирована морфометрика и её вариации одного из уникальных видов жужелиц Центральной Азии – Machozetus lehmanni. Наиболее вариабельным признаком у Machozetus lehmanni является длина головы, а наиболее стабильным признаком является ширина надкрылий. На разных участках ареала, в зависимости от почвенных условий, могут наблюдаться морфологические модификации, выражающиеся прежде всего в размерах головы.
Библиографическая ссылка
Зокирова Д.Ф., Алимова Л.Х., Халимов Ф.З. ИЗУЧЕНИЕ МОРФОМЕТРИЧЕСКИХ ОСОБЕННОСТЕЙ MACHOZETUS LEHMANNI MENETRIES, 1848 – ЭНДЕМИКА СРЕДНЕЙ АЗИИ // Научное обозрение. Биологические науки. 2022. № 1. С. 16-21;URL: https://science-biology.ru/ru/article/view?id=1253 (дата обращения: 28.07.2026).
DOI: https://doi.org/10.17513/srbs.1253