

 science-review.ru
science-review.ru
Введение
Водные клопы (отряд Hemiptera, подотряд Heteroptera) представляют собой ключевую группу насекомых, обитающих в пресноводных экосистемах. Они широко распространены в реках, озёрах, каналах, прудах, а также во временных водоёмах. Благодаря высокой численности и видовому разнообразию водные клопы играют важную роль в структуре и функционировании гидробиоценозов [1-3]. На международном уровне водомерки рассматриваются не только с биологической и экологической точки зрения, но и как важные биоиндикаторы, позволяющие оценивать состояние водных ресурсов, мелиорацию земель и физико-химические параметры воды [4; 5]. Распространение водных клопов зависит от типа водоёма, климатических условий, растительного покрова и антропогенного воздействия. За последние десятилетия проведено множество исследований, посвящённых систематике, экологии и биогеографии водных клопов [6-8].
Таблица 1
Объекты исследования водяных клопов в разных районах Хорезмской области в 2023-2025 гг. (места отбора проб)
|
№ |
Наименование объекта |
Территория |
Координаты |
|
1 |
Крупный канал Амударьи |
Ургенчский район |
41.5540° с.ш., 60.6190° в.д. |
|
2 |
Канал Палван |
Шаватский район |
41.4300° с.ш., 60.4300° в.д. |
|
3 |
Озеро Дауткёль |
Гурленский район |
41.2900° с.ш., 60.3800° в.д. |
|
4 |
Оросительный канал |
Янгибазарский район |
41.5000° с.ш., 60.7500° в.д. |
|
5 |
Пруд (центр Хорезма) |
Хивинский район |
41.3800° с.ш., 60.3600° в.д. |
|
6 |
Прибрежная зона |
Лалмикорская заболоченная территория |
41.2100° с.ш., 60.8200° в.д. |
Результаты показывают значительные изменения их ареалов, связанные с климатическими изменениями, истощением водных ресурсов, антропогенной нагрузкой и деградацией водных экосистем [9; 10].
В Центральной Азии, включая Узбекистан, исследования водных клопов начались в конце XIX века и получили активное развитие в XX веке [11]. Тем не менее имеющиеся публикации содержат лишь фрагментарные данные о распространении этих насекомых, при этом большинство видов зарегистрировано лишь в отдельных районах страны.
Цель данного исследования – изучение влияния химического состава воды на видовое разнообразие водных клопов в Хорезме. Результаты позволят выявить закономерности их распространения, экологические предпочтения и роль водомерок в локальных пресноводных экосистемах.
Материалы и методы исследования
Учеты проводились с марта до сентября. Изучение водных клопов нами проводилось в некоторых объектах в 2023-2025 гг. (табл. 1).
В рамках данного исследования применялись следующие основные методы: сбор водных клопов с помощью сачков и гидробиологических пробоотборников; определение видового состава по морфологическим признакам; количественный учёт, заключающийся в подсчёте особей на единицу площади или объёма; а также анализ сезонной динамики численности [12; 13].
Исследования проводились с учётом сезонных этапов развития водных клопов. Весной (март–апрель) насекомые пробуждаются от зимней диапаузы. В этот период отмечается появление первых имаго и личинок, начинается размножение. Весенний этап является важным для формирования годовой серии наблюдений. Физико-химические параметры воды (температура, pH, содержание кислорода) фиксировались с использованием портативных приборов [14-16].
Обзор научно-исследовательских публикаций и материалов о водных клопах, об изучении фаунистического и зоогеографического анализа в районах Хорезма позволяет сделать вывод о том, что водные полужесткокрылые в этих регионах изучены недостаточно [17; 18].
Несмотря на важное экономическое и экологическое значение водных клопов, их видовой состав, биоэкологические особенности по отдельным видам и их географическое распространение по регионам Узбекистана, а также роль клопов в природных водных экосистемах Республики до сих пор остаются малоизученными [19; 20].
Результаты исследования и их обсуждение
По результатам исследований фауна водомерок в Хорезмской области делится на три основных инфраотряда: Gerromorpha, Nepomorpha и Leptopodomorpha. В этих инфраотрядах идентифицировано в общей сложности 9 надсемейств, 11 семейств и 13 подсемейств. Водомерки относятся к 18 родам и 12 подродам, всего зарегистрировано 35 основных видов и 9 подвидов. Эти данные свидетельствуют о таксономическом разнообразии и биологическом богатстве фауны водомерок в Хорезмском оазисе.
1. GERROMORPHA Popov, 1971
• Gerridae Leach, 1815, семейство
- Aquarius paludum paludum (Fabricius, 1794)
- Limnoporus rufoscutellatus (Latreille, 1807)
- Gerris (Gerris) argentatus (Schummel, 1832)
- Gerris (Gerris) costae (Herrich-Schaeffer, 1850) и 3 подвида
- Gerris (Gerris) lacustris (Linnaeus, 1758)
- Gerris (Gerris) thoracicus (Schummel, 1832)
- Gerris (Gerris) odontogaster (Zetterstedt, 1828)
- Gerris (Gerriselloides) lateralis (Schummel, 1832)
- Heterobates dohrandti (Bianchi, 1896)
• Veliidae Brullé, 1836, семейство
- Microvelia (Microvelia) buenoi (Drake, 1920)
- Microvelia (Microvelia) reticulata (Burmeister, 1835)
- Microvelia (Picaultia) pygmaea (Dufour, 1833)
- Velia (Plesiovelia) affinis affinis (Kolenati, 1857)
• Hebridae Amyot & Serville, 1843, семейство
- Hebrus (Hebrus) kiritshenkoi (Kanyukova, 1997)
- Hebrus (Hebrus) oxianus (Kanyukova, 1997)
- Hebrus (Hebrus) pilipes (Kanyukova, 1997)
- Hebrus (Hebrus) pusillus pusillus (Fallén, 1807)
- Hebrus (Hebrusella) ruficeps (Thomson, 1871)
• Hydrometridae Billberg, 1820, семейство
- Hydrometra stagnorum (Linnaeus, 1758)
- Hydrometra gracilenta (Horváth, 1899)
• Mesoveliidae Douglas & Scott, 1867, oilasi
- Mesovelia thermalis (Horváth, 1915)
- Mesovelia furcata (Mulsant & Rey, 1852)
2. NEPOMORPHA Popov, 1968
• Nepidae Latreille, 1802, семейство
- Nepa cinerea (Linnaeus, 1758)
- Ranatra (Ranatra) linearis (Linnaeus, 1758)
• Notonectidae Latreille, 1802, семейство
- Notonecta (Notonecta) glauca glauca (Linnaeus, 1758)
• Corixidae Leach, 1815, семейство
- Corixa dentipes (Thomson, 1869)
- Sigara (Sigara) assimilis (Fieber, 1848)
- Sigara (Pseudovermicorixa) nigrolineata nigrolineata (Fieber, 1848)
• Micronectidae Jaczewski, 1924, семейство
- Micronecta (Micronecta) minutissima (Linnaeus, 1758)
• Naucoridae Leach, 1815, семейство
- Ilyocoris cimicoides cimicoides (Linnaeus, 1758)
3. LEPTOPODOMORPHA
• Saldidae Amyot & Serville, 1843, семейство
- Macrosaldula scotica (Curtis, 1835)
- Saldula melanoscela (Fieber, 1859)
- Saldula orthochila (Fieber, 1859)
Полученные результаты подтверждают возможность использования водных полужесткокрылых (Heteroptera) в качестве биоиндикаторов состояния водной среды. Изучение видового состава и численности этих насекомых имеет важное практическое значение, поскольку позволяет эффективно планировать мероприятия по контролю за вредителями. В условиях усиления антропогенной нагрузки и изменения гидрологических режимов особое значение приобретает постоянный мониторинг водных клопов как элемента системы экологического контроля (рис. 1).
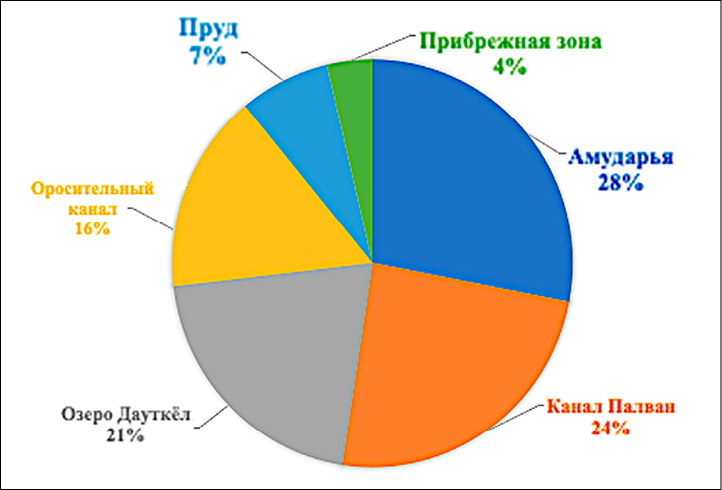
Рис. 1. Результаты мониторинга водных клопов на водоемах Хорезмской области (2023–2025 гг.) Источник: составлено авторами по результатам собственных исследований
Таблица 2
Гидрохимическая характеристика водоёмов Хорезмской области (2023–2025 гг.)
|
Водоём/ показатель |
Температура, °C |
Прозрачность, см |
Минерализация, г/л |
pH |
Растворённый O₂, мг/л |
Органические вещества, мг/л |
|
Амударья |
17–28 |
35–50 |
0.5–1.2 |
7.2–8.1 |
6.5–8.0 |
3.0–5.5 |
|
Пальван канал |
18–29 |
30–45 |
0.6–1.0 |
7.3–8.0 |
5.8–7.5 |
2.5–4.8 |
|
Озеро Дауткёль |
16–27 |
25–40 |
0.4–0.9 |
7.0–7.8 |
6.2–7.8 |
2.0–4.0 |
|
Орловский оросительный канал |
18–30 |
20–35 |
0.7–1.3 |
7.4–8.2 |
5.5–7.2 |
3.2–5.0 |
|
Пруд |
17–28 |
15–30 |
0.8–1.4 |
7.5–8.3 |
5.0–6.8 |
3.5–6.0 |
|
Береговая зона |
16–26 |
10–25 |
0.3–0.8 |
7.1–7.9 |
6.0–7.0 |
1.5–3.5 |
Для изучения условий обитания гидробионтов и влияния водной среды на прибрежную энтомофауну в период полевых исследований (2023–2025 гг.) был проведен комплексный гидрохимический анализ ключевых водных объектов Хорезмской области. Анализ включал измерения в различных типах водоемов: магистральных каналах, озерах, оросительной сети и стоячих водах (табл. 2).
Гидрохимический анализ исследованных водоёмов Хорезмской области, по данным собственных наблюдений за 2023–2025 гг., показал следующие особенности:
1. Температура воды колебалась от 16 до 30 °C в зависимости от сезона и типа водоёма. Наибольшие значения отмечались в оросительных каналах и водохранилищах летом, а минимальные – в мелких стоячих водоёмах.
2. Прозрачность воды варьировала от 10 до 50 см. Наибольшая прозрачность характерна для больших рек, таких как Амударья, а минимальная – для мелких каналов и прибрежных зон.
3. Минерализация составила 0.3–1.4 г/л. Наиболее пресной была вода в прибрежных зонах и озёрах, более солёной – в оросительных каналах и прудах, что связано с интенсивным испарением и поступлением минеральных солей.
4. pH воды колебался от 7.0 до 8.3, что соответствует слабощелочной реакции. В целом водоёмы характеризуются стабильным рН, благоприятным для большинства пресноводных организмов.
5. Содержание растворённого кислорода в воде находилось в диапазоне 5.0–8.0 мг/л. Более высокие показатели зафиксированы в проточных водоёмах, а в стоячих водоёмах и каналах кислородное насыщение было ниже, что связано с биохимическим потреблением кислорода.
6. Органическое вещество колебалось от 1.5 до 6.0 мг/л, что указывает на умеренное органическое загрязнение. Высшие значения отмечены в прудах и мелких каналах с низким течением.
В целом, гидрохимическая характеристика водоёмов Хорезмской области за 2023–2025 гг. свидетельствует об умеренно минерализованных водах с нейтрально-щелочной реакцией и достаточным кислородным насыщением, что создаёт благоприятные условия для водной флоры и фауны региона.
По данным определено, что представитель семейства Notonectidae чувствителен к снижению содержания растворённого кислорода и повышенной минерализации воды. Высокая численность N. glauca указывает на удовлетворительное качество воды с достаточным кислородным режимом и умеренной минерализацией. Снижение численности может свидетельствовать о загрязнении или деградации водоёма. Кроме того, выявлено, что некоторые виды могут служить индикаторами экологического состояния водоёмов: Notonecta glauca предпочитает чистые, слабо заросшие водоёмы; представители родов Corixa и Sigara устойчивы к умеренному уровню загрязнения; Hydrometra характерны для водоёмов с развитой прибрежной растительностью. Исследования также показали, что Sigara assimilis относительно устойчив к изменениям качества воды, но чувствителен к органическому загрязнению и эвтрофикации. Наблюдения показали, что присутствие S. assimilis в водоёме может указывать на умеренно загрязнённые или богатые органикой водоёмы. В таблице 3 выделены четыре вида из 35 зарегистрированных как биоиндикаторы. Ниже представлен их видовой состав и предпочитаемая среда обитания.
Таблица 3
Водные клопы – индикаторы состояния водной среды
|
Вид |
Предпочитаемая среда |
На что может указывать |
|
Notonecta glauca |
Чистые и умеренно загрязнённые воды |
Хорошее кислородное состояние |
|
Sigara assimilis |
Каналы и сточные воды |
Высокая органическая нагрузка |
|
Corixa dentipes |
Загрязнённые и стоячие водоёмы |
Устойчивость к эвтрофикации |
|
Gerris lacustris |
Чистые поверхностные воды |
Относительная стабильность экосистемы |
Источник: по результатам собственных исследований.
Температурный режим водоёма является одним из ключевых факторов, определяющих жизнедеятельность водных полужесткокрылых (Heteroptera). Для представителей семейств Notonectidae (плавтусы, или спиноплавы) и Gerridae (водомерки) температура воды влияет на метаболизм, скорость развития, распределение и сезонную динамику численности. Температура, содержание кислорода и минерализация воды являются ключевыми факторами, влияющими на развитие водных полужесткокрылых в Хорезме. Для проведения сравнительного анализа влияния температуры воды на водных клопов было осуществлено исследование с участием представителей двух семейств – Notonectidae и Gerridae. Проведённые исследования влияния температуры воды на водных клопов показали, что температурный фактор оказывает определяющее воздействие на их активность, численность и пространственное распределение (рис. 2).
При низких температурах воды (10–15 °C) отмечается общее снижение физиологической и трофической активности водных полужесткокрылых. Так, представители семейства Notonectidae становятся малоподвижными и значительно реже проявляют охотничье поведение. Аналогичная тенденция наблюдается у Gerridae, численность которых в холодных водоёмах существенно ограниченна (рис. 2).
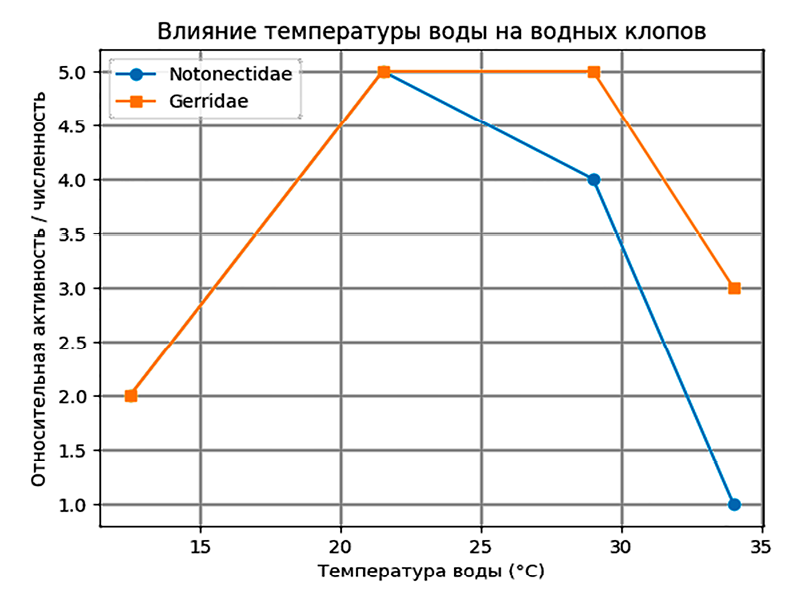
Рис. 2. Влияние температуры воды на два семейства – Notonectidae и Gerridae Источник: составлено авторами по результатам собственных исследований
Умеренный температурный диапазон (18–25°C) формирует оптимальные условия для развития большинства видов. В этих условиях у Notonectidae ускоряется развитие личиночных стадий, повышается интенсивность охоты и наблюдается рост численности популяций. Представители семейства Gerridae при данной температуре достигают максимальной численности, особенно в мелких, хорошо прогреваемых водоёмах и оросительных каналах (рис. 2).
При повышенной температуре воды (26–32 °C) Notonectidae сохраняют высокую активность, однако при длительном воздействии температур выше 30 °C отмечается снижение их численности, что связано с уменьшением содержания растворённого кислорода и усилением межвидовой конкуренции. В то же время Gerridae демонстрируют сравнительно высокую устойчивость к повышенным температурам, сохраняя значительную численность в тёплых, солнечных и слабопроточных водоёмах (рис. 2).
Критически высокие температуры воды (выше 33 °C), характерные для летнего периода, оказывают отрицательное влияние на численность водных клопов. У представителей Notonectidae наблюдается резкое сокращение популяций, особенно в пересыхающих и перегретых водоёмах. Gerridae способны выживать при таких условиях более продолжительное время, однако также снижают активность и смещаются в более глубокие или затенённые участки водоёмов (рис. 2). Сравнительный анализ численности этих семейств позволяет оценивать температурный режим и уровень устойчивости экосистемы (табл. 4).
Notonectidae могут быть индикаторами умеренно тёплых, хорошо прогретых, но не перегретых водоёмов. Gerridae показывают большую устойчивость и способны доминировать в условиях высокой температуры и сезонного колебания уровня воды. Например, при 25–28 °C ускоряется рост личинок Notonectidae и Gerridae, тогда как при температурах выше 30 °C увеличивается их смертность.
Таблица 4
Влияние температуры воды на численность Notonectidae и Gerridae
|
Температура (°C) |
Численность Notonectidae |
Численность Gerridae |
|
15 |
40 |
30 |
|
20 |
70 |
50 |
|
25 |
120 |
90 |
|
28 |
150 |
110 |
|
30 |
100 |
80 |
|
32 |
60 |
40 |
Источник: по результатам собственных исследований.
Таблица 5
Основные параметры воды и их влияние на водных клопов
|
Показатель |
Влияние на водных клопов |
Чувствительные/ устойчивые виды |
|
Кислород (O₂) |
Недостаток кислорода снижает активность и выживаемость |
Notonecta glauca чувствительны |
|
pH |
Оптимум 6.5–8.5; при отклонении – угнетение метаболизма |
Corixa, Sigara устойчивы к слабощелочной среде |
|
Минерализация (солёность) |
Умеренная солёность допустима для некоторых видов |
Gerris, Hydrometra переносят солоноватые воды |
|
Нитраты, аммиак |
Токсичны в высоких концентрациях, вызывают гибель личинок |
Nepa, Notonecta чувствительны |
|
Органическое загрязнение (БПК) |
При высоком содержании органики – изменение структуры сообществ |
Sigara assimilis и Corixa dentipes устойчивы |
|
Температура |
Повышение ускоряет развитие, но при перегреве – летальный исход |
Все виды имеют температурные пределы активности |
Источник: по результатам собственных исследований.
В рамках исследования проводился анализ влияния химических показателей воды на водных клопов, что позволило выявить их роль в распределении видов и оценить экологические условия водоёмов региона. Nepidae, обитающие в заиленных и застойных водоёмах, особенно чувствительны к уровню кислорода. Высокая минерализация воды снижает численность Nepidae. Ниже представлены данные об устойчивости водных клопов к различным условиям воды (табл. 5).
Влияние содержания растворённого кислорода в воде имеет выраженную специфику для различных семейств водных клопов. Полученные данные свидетельствуют о том, что высокое содержание кислорода (6–10 мг/л) является благоприятным фактором для представителей семейства Naucoridae, у которых отмечаются высокая двигательная и трофическая активность, увеличение численности популяций, а также успешная охота на личинок рыб и амфибий. Представители семейства Nepidae также встречаются при данных условиях, однако их численность в большей мере определяется степенью развития водной растительности и наличием подходящего субстрата, а не уровнем аэрации воды.
При среднем содержании растворённого кислорода (3–5 мг/л) Naucoridae сохраняют относительно стабильные популяции, хотя наблюдается некоторое снижение активности. В то же время Nepidae регистрируются чаще, поскольку обладают способностью выживать в условиях умеренной аэрации благодаря использованию атмосферного воздуха для дыхания.
Низкое содержание растворённого кислорода (менее 3 мг/л) оказывает негативное воздействие на представителей семейства Naucoridae, численность которых резко сокращается вследствие снижения охотничьей активности и уменьшения выживаемости личиночных стадий. В отличие от них, Nepidae демонстрируют высокую устойчивость к гипоксическим условиям, поскольку их дыхание практически не зависит от концентрации кислорода в воде. Это позволяет им заселять заиленные и эвтрофированные водоёмы.
Минерализация воды является одним из ключевых гидрохимических факторов, определяющих условия существования гидробионтов. Для водных полужесткокрылых (Heteroptera), в частности представителей семейств Gerridae (водомерки) и Nepidae (водяные скорпионы), уровень минерализации оказывает существенное влияние на их пространственное распределение, численность и эколого-физиологические особенности. Представители Gerridae обитают преимущественно на поверхности воды и отличаются высокой чувствительностью к изменениям поверхностного натяжения, солевого состава и pH среды. Nepidae, напротив, ведут преимущественно донный образ жизни, предпочитая стоячие или слаботекущие водоёмы с развитой водной растительностью, и используют дыхательный сифон для доступа к атмосферному воздуху (рис. 3).
При низкой минерализации воды (0,2–0,5 г/л) формируются оптимальные условия для большинства видов Gerridae. В таких водоёмах отмечается их высокая численность, особенно в пресных реках, каналах и озёрах. Представители семейства Nepidae при данных условиях встречаются реже, поскольку предпочитают биотопы с более высоким содержанием органического вещества.

Рис. 3. Влияние температуры воды на два семейства – Napidae и Gerridae Источник: составлено авторами по результатам собственных исследований
Средний уровень минерализации (1–3 г/л) характеризуется сохранением относительно устойчивых популяций Gerridae, однако их численность постепенно снижается. Для Nepidae такие условия являются благоприятными: они чаще заселяют водоёмы с умеренной минерализацией и повышенной трофностью, где обеспечены подходящие условия для укрытия и охоты.
Высокая минерализация воды (5–10 г/л и более) оказывает отрицательное воздействие на представителей Gerridae, которые при таких условиях практически исчезают. Это связано с нарушением поверхностного натяжения воды и осмотического баланса, что затрудняет их нормальную жизнедеятельность. Nepidae в водоёмах с высокой минерализацией встречаются единично, однако отдельные виды проявляют сравнительно высокую устойчивость благодаря донному образу жизни и способности дышать атмосферным воздухом.
На рисунке 3 отражена зависимость численности представителей семейств Gerridae и Nepidae от уровня минерализации воды. При низкой минерализации (0,2–0,5 г/л) отмечается высокая численность Gerridae, достигающая максимальных значений, что свидетельствует об оптимальных условиях для их обитания. С увеличением минерализации до 1,0–2,0 г/л наблюдается резкое снижение численности Gerridae, что связано с нарушением поверхностного натяжения воды и осмотического баланса. Численность Nepidae во всём диапазоне минерализации ниже по сравнению с Gerridae, однако снижение происходит более постепенно. Это указывает на относительную устойчивость представителей данного семейства к повышенной минерализации, обусловленную донным образом жизни и использованием атмосферного воздуха для дыхания. В целом результаты подтверждают, что Gerridae являются более чувствительными к изменениям минерализации воды и могут рассматриваться как индикаторы пресных водоёмов, тогда как Nepidae демонстрируют более широкую экологическую пластичность.
Густота водной растительности, глубина водоёма, температурный режим воды, а также степень её загрязнения напрямую влияют на численность и видовое разнообразие водных насекомых. Фауна водных полужесткокрылых Хорезмской области отличается высоким уровнем разнообразия и включает как широко распространённые, так и локально встречающиеся виды. По результатам учётов установлено, что в стоячих водоёмах преобладали Notonecta glauca, Sigara lateralis и Hydrometra stagnorum, тогда как в проточных водоёмах (каналах и арыках) наиболее часто отмечались Gerris lacustris и Corixa dentipes. В застойных мелководных участках были зарегистрированы такие виды, как Nepa cinerea и Ranatra linearis.
Заключение
Изучение водных клопов имеет важное значение для гидробиологии, экологии и природоохранной деятельности. Эти насекомые могут использоваться как биоиндикаторы состояния водоёмов и одновременно как естественные регуляторы численности насекомых-вредителей. В условиях изменения климата и антропогенного давления становятся особенно актуальными исследования их биоразнообразия и экологии.
Вода является ключевым лимитирующим фактором для водных полужесткокрылых. Температура, содержание кислорода, минерализация и колебания уровня воды оказывают прямое влияние на их численность, распределение и жизненный цикл. Благодаря высокой чувствительности к изменениям качества воды водные полужесткокрылые могут использоваться как индикаторы состояния пресноводных экосистем. Полученные данные подтверждают необходимость комплексного изучения этой группы для мониторинга водных биоценозов.
Состав воды напрямую влияет на структуру и динамику сообществ водных полужесткокрылых. Воды с низким загрязнением и стабильным кислородным режимом обогащены разнообразием видов. В эвтрофных и сточных водоёмах доминируют устойчивые, малочувствительные формы. Таким образом, водные клопы могут выступать как биоиндикаторы качества воды и использоваться в экологическом мониторинге.
Конфликт интересов
Финансирование
Библиографическая ссылка
Беглиев C., Ганджаева Л. ВЛИЯНИЕ ХИМИЧЕСКОГО СОСТАВА ВОДЫ НА ВОДНЫХ ПОЛУЖЕСТКОКРЫЛЫХ (HETEROPTERA) // Научное обозрение. Биологические науки. 2026. № 2. С. 5-13;URL: https://science-biology.ru/ru/article/view?id=1454 (дата обращения: 28.07.2026).
DOI: https://doi.org/10.17513/srbs.1454