

 science-review.ru
science-review.ru
Scientific journal
Scientific Review. Biological science
ISSN 2500-3399
ПИ №ФС77-57454
INFLUENCE OF THE CHRONIC IRRADIATION AND WEATHER CONDITIONS ON POPULATIONS OF THE SCOTS PINE GROWING IN TERRITORIES, HAVE UNDERGONE TO RADIOACTIVE POLLUTION

В настоящее время общепризнано, что система нормирования радиационного воздействия на биоту должна ориентироваться на защиту популяций [18]. Для эффективного нормирования антропогенного воздействия на популяционном уровне необходимо учитывать, что популяция это сложная иерархическая система, в которой животные и растения испытывают воздействие комплекса естественных и антропогенных факторов. Поэтому для понимания степени опасности того или иного фактора необходимо представлять как он влияет на живые организмы во взаимодействии с другими факторами окружающей среды, как эти влияния проявляются на разных уровнях организации живого, какова динамика вызываемых им биологических процессов. Для этого необходимы комплексные, долговременные наблюдения за растениями и животными в естественных условиях их обитания, однако подобных исследований в настоящее время крайне мало. Это порождает острый дефицит информации о радиационно-индуцированных эффектах у представителей живой природы в условиях хронического облучения в естественной среде их обитания [10, 11]. Особенно актуальны такие исследования для лесов на территориях, загрязненных радионуклидами в результате аварии на Чернобыльской АЭС. Только в Российской Федерации радиоактивному загрязнению подверглось около 1 млн. гектаров земель лесного фонда [13]. Особую важность такого рода исследованиям придает тот факт, что хроническое техногенное воздействие даже в низких дозах способно влиять на такие важные для существования популяции параметры как состояние и репродуктивная способность составляющих ее индивидов [5, 19, 20, 21].
Наша лаборатория с 2003 года по настоящее время проводит комплексное изучение биологических эффектов в популяциях сосны обыкновенной на шести участках Брянской области [15, 17]. Целью настоящей работы является оценка цитогенетических эффектов и репродуктивной способности популяций сосны обыкновенной в отдалённый период после аварии на Чернобыльской АЭС.
Материалы и методы
На протяжении двенадцати лет исследовали шесть популяций сосны (Pinus sylvestris L.) из Брянской области. В качестве тест объекта сосна обыкновенная была выбрана потому, что это основной лесообразующий вид эдификатор Северной Евразии. Обладая высокой радиочувствительностью, она стала одним из референтных биологических видов, на которых базируется современная концепция радиационной защиты окружающей среды [18]. К воздействию ионизирующих излучений у сосны наиболее чувствительны репродуктивные органы, отличающиеся сложностью организации и длительностью генеративного цикла (с момента закладки примордиев генеративных органов до созревания семян проходит 28 месяцев [7]). В условиях хронического действия техногенных факторов столь длительный цикл развития ведёт к накоплению в неспециализированных инициальных клетках семян достаточного для индикации внешнего воздействия количества повреждений ДНК, реализация которых в аберрации происходит главным образом в первом митозе [16].
Исследовали популяции, произрастающие на контрольных участках: К; К1 и загрязнённых радионуклидами участках, находящиеся в 200-250 км от Чернобыльской АЭС: ВИУА (ВИУА); Старые Бобовичи (СБ); Заборье поле (ЗП); Заборье кладбище (ЗК). Все участки характеризуются высоким представительством сосновых деревьев в фитоценозе, однородностью физико-химических свойств почв, а также уровнем техногенного загрязнения (рис. 1).
На экспериментальных участках брали пробы почв и собирали шишки. Образцы почвы на каждом участке отбирали по слоям 0-5 см, 5-10 см, 10-15 см в нескольких местах под кроной деревьев, где была зафиксирована максимальная на участке мощность дозы. Почву высушивали до воздушно-сухого состояния, измельчали и просеивали через сито с размером отверстий 1 мм. Шишки собирали с деревьев 30-50-летнего возраста в ноябре – начале декабря 2003-2014 гг. На каждом участке шишки собирали с 20-29 деревьев в пределах гомогенного древостоя по 20-50 шишек с каждого дерева на высоте 1.5-2 м от поверхности земли. Для дозревания и стратификации шишки выдерживали вне помещения до конца февраля. Затем их доставляли в лабораторию и хранили при комнатной температуре и низкой влажности до раскрытия и высыпания семян, которые обескрыливали вручную. Для исследований использовали только свободно высыпавшиеся, хорошо выполненные семена.
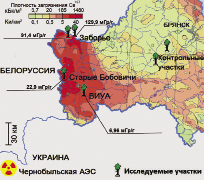
Рис. 1. Расположение экспериментальных участков
В образцах почвы и шишках определяли активность 137Cs, методом g-спектрометрии на многоканальном анализаторе IN-1200 (Франция) с германиевым детектором GEM-1200 (США). Также в шишках определяли содержание 90Sr радиохимическим методом.
Данные по метеоусловиям на экспериментальных участках были предоставлены ВНИИ гидрометеорологической информации – Мировым Центром Данных (ВНИИГМИ МЦД, г. Обнинск). Использовали данные четырех ближайших к экспериментальным участкам метеостанций, расположенных в населенных пунктах Брянск (К), Унеча (К1), Трубчевск (ВИУА и СБ) и Красная Горка (ЗП и ЗК).
Для оценки репродуктивной способности визуально определяли число нормальных и абортивных (недоразвитые, пустые, сухие) семян на шишку (урожай 2007-2014 гг.). При проращивании семян урожая 2008-2014 гг. оценивали их всхожесть (как процент нормально развитых проростков на 20-тый день). Перед проращиванием, для синхронизации деления, семена выдерживали сутки в холодильнике при температуре 4°С, затем проращивали в термостате при 24°С в чашках Петри на смоченной дистиллированной водой фильтровальной бумаге. Эксперимент проводили в 4 повторностях, по 50 семян в каждой.
Для цитогенетического анализа использовали проростки семян с корешками длиной 7-10 миллиметров, когда наблюдается пик первых митозов [16]. Корешки фиксировали в ацето-алкоголе (1:3), окрашивали ацетоорсеином и готовили временные давленые препараты. Все препараты кодировали. В каждом из 20-100 препаратов анализировали все ана-телофазные клетки (4400-14000 ана-телофаз на вариант опыта) и рассчитывали долю клеток с цитогенетическими нарушениями. При анализе спектра нарушений выделяли хроматидные (одиночные) и хромосомные (двойные) мосты и фрагменты, многополюсные митозы, а также отставания хромосом. Отметим, что анафазным методом в клетках корневой меристемы проростков регистрируются нарушения, возникшие в период от образования гамет до созревания и сбора семян, поскольку индуцированные на вегетативной стадии (до цветения) перестройки хромосом элиминируются в мейозе за исключением не регистрируемых этим методом симметричных транслокаций и инверсий.
Часть собранных в 2003, 2004, 2006 и 2013гг. семян подвергли острому γ-облучению в дозе 15 Гр с мощностью дозы 0,6 Гр/мин от источника 60Сo (установка «Луч», Латвия). Сразу после облучения семена проращивали и определяли частоту цитогенетических нарушений в корневой меристеме проростков. В зависимости от года и участка анализировали от 10 до 24 препаратов (1000-4500 ана-телофаз) на вариант опыта.
Экспериментальные данные проверяли на наличие выбросов по критерию Диксона. Выбросы исключали из дальнейшего рассмотрения. Данные обрабатывали методами вариационной статистики с использованием MS Excel. Для оптимизации объема выборки применяли методику статистического анализа эмпирических распределений [4]. Достоверность отличий оценивали с помощью критерия Стьюдента.
Результаты исследования
Почвы всех исследованных участков близки по физико-химическим свойствам, а содержание тяжелых металлов и доступных растениям их подвижных форм не превышает допустимых уровней [3, 17]. Качественно иная картина имеет место в отношении радиоактивного загрязнения участков, основной вклад в которое вносит 137Cs – ведущий дозообразующий радионуклид на загрязненных в результате аварии на Чернобыльской АЭС территориях. Поскольку период полураспада 137Cs составляет 30 лет, радиоактивное загрязнение участков за период исследования изменилось незначительно. Поэтому в табл. 1 представлены данные о радиоактивном загрязнении участков за один конкретный (2013) год.
Содержание 137Cs в верхнем 5-см слое почвы изменяется в широком диапазоне и на импактных участках значительно выше, чем в контроле. Отметим, что удельная активность 137Cs в почвах контрольных участков К и К1 различается в два раза, однако эти колебания лежат в пределах, характерных для региона, и обусловлены глобальными выпадениями 137Cs [2].
Содержание радионуклидов в шишках деревьев импактных участков статистически значимо превышает этот параметр для контрольных популяций (табл. 1). Причем содержание 137Cs в шишках во всех случаях выше содержания 90Sr как минимум на порядок. Максимальные удельные активности 137Cs и 90Sr в шишках импактных участков существенно превышают аналогичные показатели для контрольных участков.
Таблица 1
Удельная активность радионуклидов и дозы (внешнее и внутреннее облучение) поглощённые короной сосны (включая естественный радиационный фон).
|
Участок |
Удельная активность, Бк/кг |
Доза, поглощенная кроной в 2013 году, мГр/г. |
Доза, поглощенная кроной за период формирования семян (2012-2013 год), мГр |
||
|
шишки |
Почва (верхний 5-см слой) |
||||
|
137Cs |
90Sr |
137Cs |
|||
|
К |
4,2 |
1,12 |
13,2 |
0.90 |
1.81 |
|
К1 |
12,6 |
0,79 |
156 |
1.11 |
2.22 |
|
ВИУА |
207 |
11,34 |
10800 |
10.9 |
21.8 |
|
СБ |
302 |
35,91 |
13000 |
20.3 |
40.6 |
|
ЗП |
2170 |
43,22 |
35600 |
34.0 |
68.0 |
|
ЗК |
1420 |
48,69 |
46200 |
39.5 |
79.0 |
Для оценки дозовых нагрузок на крону и генеративные органы изучаемых популяций сосны от внешнего и внутреннего облучения, была разработана дозиметрическая модель, подробное описание которой дано в [17]. Согласно расчетам, дозовая нагрузка на кроны сосновых деревьев находится в диапазоне 10.9-39.5 мГр/г. (табл. 1).
В популяциях, населяющих загрязнённые радионуклидами участки, частота цитогенетических нарушений статистически значимо превышает (рис. 2) соответствующие контрольные значения на протяжении всех лет исследования (2003–2014). Максимальная частота аберрантных клеток зафиксирована в проростках с участков ЗП и ЗК, характеризующихся наибольшими значениями поглощенных репродуктивными органами сосны доз.
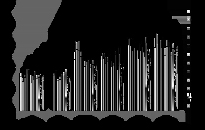
Рис. 2. Частота цитогенетических нарушений в корневой меристеме проростков семян. * - отличие от к и к1 значимо
Более того, на протяжении практически всех лет исследования наблюдалась статистически значимая корреляция (r=0.70–0.99; p<1%) между частотой аберрантных клеток в проростках семян и мощностью экспозиционной дозы на участке.
Частота цитогенетических нарушений в 2006–2014 гг. статистически значимо возрастала с увеличением поглощенной дозы (r = 0.81–0.94; p < 0.05), а в 2007–2011 гг. и удельной активности 137Cs в верхнем слое почвы (r = 0.77–0.89; p < 0.05). В 2003–2005 и 2014 гг. частота цитогенетических нарушений также увеличивалась с дозой, однако эта зависимость не была статистически значимой.
Анализ спектра цитогенетических нарушений показал, что в популяциях сосны, населяющих наиболее загрязненные радионуклидами участки – ЗП и ЗК – частота маркеров радиационного воздействия (аберраций хромосомного типа) и митотических аномалий (отставаний хромосом и многополюсных митозов) значительно превышала контрольный уровень на протяжении всех лет исследования. Причем в большинстве случаев это различие было статистически значимым.
При длительном воздействии неблагоприятного фактора популяция может к нему адаптироваться. Выявить наличие адаптации к действию радиационного фактора можно при помощи дополнительного облучения высокой дозой [6, 12, 14]. Часть собранных в 2003, 2004, 2006 и 2013 гг. семян перед проращиванием была подвергнута острому g-облучению. Полученные результаты свидетельствуют об отсутствии увеличения радиоустойчивости в популяциях сосны, длительное время развивавшихся в условиях низкодозового хронического облучения (рис. 3).
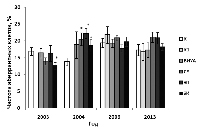
Рис. 3. Частота цитогенетических нарушений в корневой меристеме проростков семян подвергшихся дополнительному острому g-облучению. * - отличие от к и к1 значимо
Эффект радиоадаптации в природных популяциях растений неоднократно был показан в исследованиях на ВУРСе и в 30-км зоне ЧАЭС [11, 14]. В том числе, и на семенах сосны. В тоже время, есть немало примеров, когда повышение радиоустойчивости в подвергающихся хроническому воздействию популяциях растений не происходит [1, 10]. Таким образом, скорость и сама возможность формирования этого признака могут существенно различаться в разных экологических условиях.
При анализе представленных данных возникает вопрос: как выявленный в нашем исследовании повышенный уровень мутагенеза в популяциях сосны, развивающихся в условиях хронического облучения, проявляется на более высоких уровнях организации живого, в том числе на показателях жизнеспособности? Анализ качества семян не выявил взаимосвязи между долей абортивных семян и величиной радиационной нагрузки. Всхожесть семян, хотя и характеризовалась значительно большим разбросом значений, что проявилось в наличии статистически значимых отличий от контроля, также не проявляла зависимости от величины радиационного воздействия (р>5%) (рис. 4 и 5).
Поскольку нам не удалось обнаружить связи качества семян с поглощенными ими дозами, а загрязнение участков тяжелыми металлами не превышает допустимых уровней [17], логично предположить, что наблюдаемая изменчивость изучаемых параметров определяется другими факторами. В отдаленный период после радиационных аварий, когда, вследствие радиоактивного распада, радиационная нагрузка на растения существенно снизилась, на первый план по силе влияния выходят иные факторы, среди которых важное место занимают метеорологические условия в период формирования семян [8, 9].
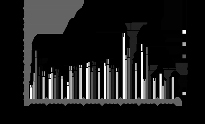
Рис. 4. Количество абортивных семян.
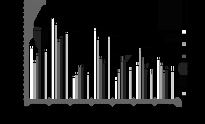
Рис. 5. Всхожесть семян. * - отличие от к и к1 значимо
Действительно, анализ зависимости показателей качества семян от погодных условий выявил в ряде случаев наличие статистически значимых корреляций (табл. 2), согласно которым повышенные температуры в течение всего периода развития семян и в августе второго года их развития снижают всхожесть, а в мае первого года (когда происходит опыление) снижают количество абортивных семян. Большое количество осадков, напротив, снижает всхожесть семян при воздействии в течение всего периода их развития, а в мае первого года развития увеличивает количество абортивных семян. Отметим, что, в отличие от представленных в [10] результатов, нам не удалость установить различий в реакции на погодные условия растений из хронически облучающихся и контрольных популяций. Это может быть связано с различием радиоэкологических условий на ВУРСе и чернобыльском следе, кардинальными различиями в биологии рассматривавшихся в этих работах видов растений, а также недостаточным объемом имеющихся у нас экспериментальных данных.
Таблица 2
Статистически значимые корреляции показателей качества семян с погодными условиями
|
Вариант |
Количество абортивных семян |
Всхожесть |
||
|
Σ Р |
Тav ef |
Σ Р |
Тav ef |
|
|
2 года |
||||
|
СБ |
-0,84 |
|||
|
ЗП |
-0,88 |
|||
|
ЗК |
-0,86 |
|||
|
Май первого года развития семян |
||||
|
СБ |
||||
|
ЗП |
0,77 |
-0,89 |
- |
|
|
Август второго года развития семян |
||||
|
К |
-0,80 |
|||
Тav ef – сумма эффективных температур, Σ Р – сумма осадков, D – число дней с эффективной температурой (t> 5°C).
Заключение
Таким образом, в исследованных популяциях сосны обыкновенной формируется семенное потомство с повышенным уровнем цитогенетических нарушений. Сопоставление частоты цитогенетических нарушений с характеристиками радиационной ситуации на экспериментальных участках свидетельствует об увеличении частоты мутаций с ростом радиационной нагрузки. При этом, несмотря на то, что радиационный фактор воздействует на изученные популяции уже не одно десятилетие, признаков радиоадаптации не обнаружено.
Хроническое облучение, вызывая устойчиво воспроизводившуюся у импактных популяций за время исследования (2003-2014 гг.) повышенную частоту цитогенетических нарушений не оказало существенного влияния на их репродуктивную способность. В настоящее время, спустя 30 лет после аварии на Чернобыльской АЭС, погодные условия оказывают гораздо большее влияние на качество семян, чем радиоактивное загрязнение.
Работа выполнена при поддержке Российского научного фонда (проект № 14-14-00666).
Библиографическая ссылка
Васильев Д.В., Кузьменков А.Г., Дикарева Н.С., Гераськин С.А. ВЛИЯНИЕ ХРОНИЧЕСКОГО ОБЛУЧЕНИЯ И ПОГОДНЫХ УСЛОВИЙ НА ПОПУЛЯЦИИ СОСНЫ ОБЫКНОВЕННОЙ ПРОИЗРАСТАЮЩИЕ НА ТЕРРИТОРИЯХ, ПОДВЕРГШИХСЯ РАДИОАКТИВНОМУ ЗАГРЯЗНЕНИЮ В РЕЗУЛЬТАТЕ АВАРИИ НА ЧЕРНОБЫЛЬСКОЙ АЭС // Научное обозрение. Биологические науки. 2016. № 6. С. 14-19;URL: https://science-biology.ru/en/article/view?id=1017 (дата обращения: 19.07.2026).