

 science-review.ru
science-review.ru
Scientific journal
Scientific Review. Biological science
ISSN 2500-3399
ПИ №ФС77-57454
METHOD FOR PROCACCI GRASSBERGER-DETERMINATION OF THE FRACTAL INDICATORS MORPHOLOGICAL STRUCTURES

In nonlinear mechanics the state of the system in the multidimensional space describes the phase trajectory. Which over time seek to some subset of it, called the attractor of the dynamic system. Its dimension is less than the original phase space. The structure of this subset can be extremely difficult, but self-similar at different scales. Geometric properties of such sets are characterized by the notions of fractal dimensions. In our case of interest is the correlation dimension, since it is much easier to compute compared to others, and it gives a good characterization of the complexity of the attractor. Build dependency logC(ε) logε possible from the slope of the linear section of this curve to calculate the magnitude of the correlation dimension. It is shown that the process of recovery of the ultrastructure of cilia and structure of the ependymal cells in the period of re-adaptation of rats to the horizontal position indicates, apparently, the recovery of the outflow of CSF from the ventricles. A much less pronounced destructive changes in the ependymal cells in repeated 14-day ANOP (compared to the initial 14 hours of power( evidenced by the fact that when re-ANOP, despite repeated blood redistribution in the cranial direction, the pressure increase in the ventricles of the brain was smaller than in primary ANOP, perhaps as a result of increasing the efficiency of the mechanism of CSF outflow through Pohitonova granulation. The results of the study are largely consistent with previously discovered by minimizing changes in the blood vessels of the brain of rats during delayed for a long time re modeling of the effects of weightlessness.
В нелинейной механике состояние системы в многомерном пространстве описывается фазовой траекторией. Которая с течением времени стремиться к некоторому его подмножеству, который называется аттрактором динамической системы. Его размерность меньше чем исходное фазовое пространство. Структура данного подмножества может быть чрезвычайно сложной, но самоподобной на различных масштабах. Геометрические свойств таких множеств характеризуются понятиями фрактальных размерностей. В нашем случае представляет интерес корреляционная размерность, поскольку её значительно проще вычислять, по сравнению с другими, и она даёт хорошую характеристику сложности аттрактора. Корреляционная размерность есть предел:
logC(ε)
D= lim —
ε ->0 logε
если данный предел существует. Это утверждение основывается на теореме Такенса [1].
Построив зависимость logC(ε) от logε можно по наклону линейного участка этой кривой рассчитать величину корреляционной размерности. Для проведения этой оценки необходимо, чтобы выполнялось условие С(ε) = const•εD. Посколько временной ряд ограничен, то линейность графиков корреляционного интеграла может нарушаться на больших масштабах. На малых масштабах искажения связаны с дискретностью данных.
Табличный метод фрактального анализа (рис.1) заключается в разделении изображения на сектора ячейки с последующим подсчетом количества секторов, потребовавшихся для покрытия всего изображения.
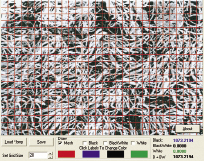
Рис 1. Табличный метод фрактального анализа
Повторение данного действия с использованием секторов разной размеренности позволят построить логарифмическую функцию зависимости размера секторов (Ось X) и количества секторов, необходимых для покрытия всего изображения (Ось Y).
Крутизна этой функции будет отображать размеренность сектора. Размеренность сектора считается максимально приближенной к фрактальной размеренности.
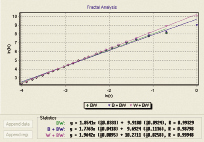
Рис 2. Графическое определение фрактальной размерности, заданной в табличной форме
Актуальность исследования заключается в возможности математического описания морфологических структур при перераспределение жидких сред в краниальном направлении у крыс, находящихся в антиортостатическом положении при вывешивании за хвост АНОП. Данный подход моделирует один из эффектов невесомости, обнаруженный у человека в космическом полете. Моделирование этого эффекта сопровождается у крыс морфологическими изменениями сосудов мозга, сосудистого сплетения и эпендимоцитов [3,6], указывающими на затруднение оттока крови от головного мозга и замедление оттока цереброспинальной жидкости (ЦСЖ) из желудочков. Повторное пребывание крыс в АНОП - несмотря на повторное перераспределение жидких сред - минимализирует изменения, возникающие в сосудистой системе мозга и в сосудистом сплетении желудочков [4,5].
Материалы и методы
Целью исследования являлось изучение механизмов адаптации организмов к длительному повторному антиортостатическому положению (АНОП), моделирующему некоторые эффекты невесомости, возникающие у человека в космическом полете.
В настоящей работе с помощью сканирующей электронной микроскопии исследовали эпендиму третьего желудочка мозга крыс, первично и повторно находившихся в условиях АНОП. В эксперименте использовали крыс самцов линии Вистар с исходной средней массой тела 172 ± 1 г., разделенных на 7 групп по 7 животных в каждой. Крысы 1-й группы являлись контролем ко 2-й группе; крысы 2-й группы были вывешены в АНОП в течение 30 суток; животные 3-й группы - контроль к 4-й группе; в 4-й группе крысы были вывешены в АНОП 30 суток и реадаптированы затем к горизонтальному положению в течение 30 суток; крысы 5-й группы - контроль к животным 6-й и 7-й групп; крысы 6-й группы были впервые вывешены в АНОП на 14 суток, а крысы 7-й группы находились в АНОП 14 суток, затем реадаптированы в течение 30 суток к горизонтальному положению и повторно были вывешены в АНОП в течение 14 суток. Для вывешивания крыс в АНОП при угле между телом и полом клетки ~ 30) использовали устройство, предложенное E.R.Morey [7] и модифицированное Е.А.Ильиным и В.Е.Новиковым [2].
Животных забивали декапитацией с помощью гильотины. Для сканирующей электронной микроскопии образцы ткани мозга, содержащие слой эпендимальных клеток, вместе с подлежащей тканью вырезали из стенки третьего желудочка вблизи сосудистого сплетения, фиксировали в 2,5 % глутаральдегиде в 0,1М какодилатном буфере рН 7,3 при 4 0С, дофиксировали в 1 % Оs О4, обезвоживали в этаноле возрастающих концентраций и в ацетоне, высушивали методом критической точки в углекислоте, монтировали с помощью серебряной краски на алюминиевые блоки, покрывали золотом в спуттере и и исследовали в сканирующем электронном микроскопе JSM - 840A при увеличении х 1000 - 5000.
Результаты и обсуждение
У крыс виварийного контроля на поверхности стенки третьего желудочка мозга видны реснички и микроворсинки эпендимальных клеток, причем реснички собраны в пучки и имеют четкую ориентацию в одном направлении (рис 3).

Рис 3. . Реснички эпедимальных клеток третьего желудочка мозга крысы виварийного контроля. Бар – 10 μm
Реснички необычайно плотно расположены и густо покрывают поверхность эпендимы таким образом, что сквозь них трудно различима поверхность эпендимальных клеток и расположенные на ней микроворсинки. Строение поверхности эпендимы совпадает с описанием строения аналогичной структуры у крыс, исследовавшейся с помощью сканирующей электронной микроскопии ранее [8,9]. Ультраструктура поверхности эпендимы у крыс виварийного контроля 1-й, 3-й и 5-й групп была одинакова.
У крыс, находившихся 30 сут. в условиях АНОП, поверхность эпендимы характеризовалась резким сниженим общего количества ресничек или полным их отсутсвием в ряде мест, а также потерей ориентации ресничек (рис. 4).
Пучки ресничек содержат меньшее их количество, реснички расположены хаотично и часто соединены между собой. На поверхности эпендимальных клеток, не покрытых ресничками, также видно резкое уменьшение числа микроворсинок или отмечается их отсутствие.

Рис 4. Реснички эпедимальных клеток третьего желудочка мозга крысы, находившейся 30 суток в условиях АНОП. Снижение численности и хаотичная ориентация. Бар – 10 μm
Пребывание крыс в течение 30 суток в горизонтальном положении после 30-суточного АНОП вызвало почти полное восстановление поверхностных структур эпендимы (рис.5).
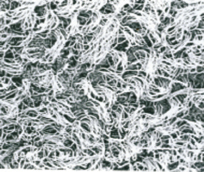
Рис 5. Реснички эпедимальных клеток третьего желудочка мозга крысы, реадаптированной 30 суток к горизонтальному положению после 30-суточного АНОП. Почти полное восстановление численности и ориентации. Бар – 10 μm
Возросла численность ресничек в пучках. Реснички приобрели более прямой вид и более четкую ориентацию. Уменьшилась площадь эпендимы не покрытая ресничками и увеличилась численность микроворсинок на поверхности эпендимальных клеток. В целом, строение поверхностных структур эпендимальных клеток приблизилось к таковому у крыс виварийного контроля, однако полного восстановления ультраструктуры не наступало. Повторное 14-суточное воздействие АНОП на крыс после их 30-суточной реабилитации к горизонтальному положению вызвало уменьшение численности ресничек на поверхности эпендимы по сранению с предшествующим воздействием реадаптации (рис.6). Уменьшилось количество ресничек в пучках и стала более неопределенной их ориентация. Однако, эти изменения ультраструктуры были значительно менее выражены по сравнению с изменениями как после 30-суточного АНОП, так и после первичного 14-суточного АНОП (рис.7). В свою очередь, изменения ультраструктуры поверхности эпендимы после первичного 14-суточного АНОП по своей выраженности занимали промежуточное положение между ультраструктурой эпендимы крыс виварийного контроля и ее изменениями после первичного 30-суточного АНОП, что свидетельствует о нарастании деструктивных изменений ультраструктуры по мере увеличения продолжительности АНОП.
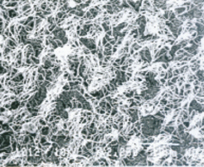
Рис. 6. Реснички и микроворсинки эпедимальных клеток 3-го желудочка мозга крысы, впервые находившейся в 14-суточном АНОП. Бар – 10 μm
Известно, что движение ресничек осуществляет перемещение ЦСЖ в пространстве желудочков и в направлении к Пахитоновым грануляциям саггитального венозного синуса, через которые осуществляется отток ЦСЖ в венозную систему. Полученные данные о резком уменьшении численности ресничек эпендимальных клеток у крыс, находившихся в 30-суточном АНОП, свидетельствует о нарушении ликвородинамики в желудочках мозга. В то же время эти данные, а также выявленное уплощение эпендимальных клеток и отек слоя в сочетании с уплощением эпителия ворсин сосудистого сплетения, обнаруженном при гистологическом исследовании мозга тех же крыс ранее [4,5], свидетельствуют о повышении внутрижелудочкового давления, являющегося, очевидно, результатом нарушения оттока ЦСЖ при перераспределении жидких сред организма в краниальном направлении.
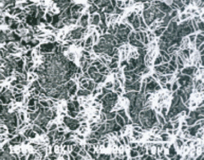
Рис. 7. Реснички и микроворсинки эпедимальных клеток 3-го желудочка мозга крысы, повторно находившейся в 14-суток в АНОП. Большая численность ресничек по сравнению с их количеством полсе первичного 14-суточного АНОП. Бар – 10 μm
Процесс восстановления ультраструктуры ресничек и строения эпендимальных клеток в период реадаптации крыс к горизонтальному положению указывает, по-видимому, на восстановление оттока ЦСЖ из желудочков. Значительно менее выраженные деструктивные изменения эпендимальных клеток при повторном 14-суточном АНОП (по сравнению с первичным 14-суточным( свидетельствует о том, что при повторном АНОП, несмотря на повторное перераспределение крови в краниальном направлении, повышение давления в желудочках мозга стало меньшим, чем при первичном АНОП, возможно, в результате повышения эффективности механизма оттока ЦСЖ через Пахитоновы грануляции. Результаты проведенного исследования в целом совпадают с обнаруженной ранее минимализацией изменений в сосудах мозга крыс при отставленном на длительное время повторном моделировании эффектов невесомости [4,5].
На рисунке 8 видно, что полученные фрактальные характеристики оптимально отражают полученные морфологические струтуры длительного антиортостатического эксперимента.
Работа выполнена при поддержке Фонда фундаментальных исследований ГНЦ РФ ИМБП.
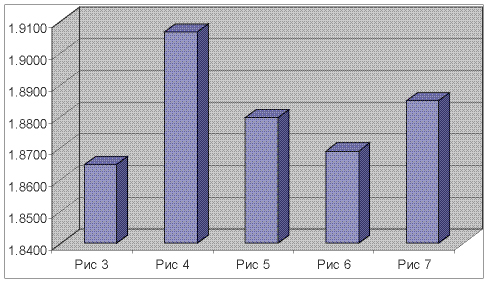
Рис 8. Фрактальные характеристики полученных морфологических структур
Библиографическая ссылка
Носовский А.М., Сидоренко Л.А. МЕТОД ГРАССБЕРГЕРА-ПРОКАЧЧИА ДЛЯ ОПРЕДЕЛЕНИЯ ФРАКТАЛЬНЫХ ПОКАЗАТЕЛЕЙ МОРФОЛОГИЧЕСКИХ СТРУКТУРЫ // Научное обозрение. Биологические науки. 2016. № 6. С. 48-52;URL: https://science-biology.ru/en/article/view?id=1022 (дата обращения: 19.07.2026).