

 science-review.ru
science-review.ru
В условиях современной жизни, когда необходимы решение вопросов экологической генетики и разработка новых тест-систем для определения потенциальной опасности новых веществ, количество которых увеличивается год от года, значительно возрос интерес к исследованиям на Drosophila melanogaster [10-13]. Drosophila melanogaster - один из наиболее распространённых модельных объектов, обладающих всеми качествами, необходимыми для успешного проведения генетического анализа: 1) незначительный срок развития от яйца до взрослой мухи (10 дней); 2) исключительно высокая плодовитость; 3) богатство наследственных рас или мутаций; 4) малое число хромосом (2n=8); 5) сходные с другими эукариотами пути метаболизма многих химических соединений [9].
Целью нашего исследования было создание линии Drosophila melanogaster со сцепленными маркерами по I хромосоме и изучение её генетических (спонтанный кроссинговер) и экологических (численность, половая пропорция) особенностей.
В экспериментах использовалась линия yw – (y – yellow (желтое тело) – I, 0.0; w – white (белые глаза) – I, 1,5 и линия дикого типа Oregon-R) [3]. В лабораторных условиях культуры дрозофилы содержали в термостате при t =24 -25°С на банановой среде. Для определения частоты спонтанного кроссинговера было поставлено 30 пробирочных культур, в которых одна виргильная самка yw/++ скрещивалось с одним самцом yw/Y. Эксперимент проводился в 5 повторностях, т.е. каждый раз родителей содержали вместе 5 дней, затем перебрасывали на новую питательную среду. В ходе исследования в потомстве вёлся учёт мух всех фенотипических классов, как среди самок, так и среди самцов. Потомство появилось только в 18 культурах. Учет дочернего поколения велся до полного вылета имаго. Частоту рекомбинации определяли по результатам анализирующего скрещивания [9]. Оценивали значимость отклонения от теоретически ожидаемого результата методом -2 [6].
Явление кроссинговера было открыто на дрозофиле Т. Х. Морганом и оказалось общим для всех животных, растений, микроорганизмов [5, 8]. Современные модели кроссинговера являются производными от модели Холидея [12]. Модель Холидея включает следующие стадии: 1) разъединение полинуклеотидных цепей двойной спирали ДНК в каждой из двух гомологичных хроматид; 2) образование «гибридных» молекул ДНК, образующихся из полинуклеотидной цепи, принадлежащей одной хроматиде, и другой полинуклеотидной цепи, принадлежащей гомологичной хроматиде; 3) генную конверсию, возникающую в результате замены ошибочно вставленных оснований в «гибридных» молекулах ДНК. Из природы кроссинговера следует, что он должен включать разрывы молекул ДНК и затем синтез ДНК, необходимый для того, чтобы соединить разорванные молекулы, которые участвуют в кроссинговере.
Таким образом, существует следующий порядок событий для мейотического кроссинговера: вначале происходит образование разрывов в специфических участках молекул ДНК с помощью соответствующей эндонуклеазы. Затем возникают одноцепочные участки ДНК. Существует белок, который облегчает реассоциацию одноцепочных участков, и в конце концов все участки молекул ДНК, вовлечённые в кроссинговер, соединяются с помощью репарирующих механизмов [13].
Для того чтобы происходил кроссинговер и формировались хиазмы, необходимо присутствие синаптонемального комплекса [5]. Если синаптонемальный комплекс отсутствует в силу внешних или внутренних причин (т.е. в результате мутаций или экспериментального вмешательства), кроссинговер не происходит и хиазмы не формируются [12].
Частота мейотического кроссинговера (rf - frequencies recombination) зависит от многих факторов окружающей среды. Различные типы излучений: ультрафиолетовый свет, рентгеновские и γ-лучи, корпускулярное излучение, как правило, повышают частоту рекомбинации, вызывая одно- и двунитевые разрывы в ДНК хромосом [13]. Кроме того, частота рекомбинации может увеличиваться в популяциях, которые периодически подвергаются стрессовым воздействиям, а также связано с возрастом и полом особи [4].
Согласно одному из постулатов биологической эволюционной аксиомы, “эволюция органического мира была бы невозможной без генетических рекомбинаций или событий, служивших их аналогами” [5]. Линейное расположение генов в хромосоме обеспечивает свободную комбинаторику генов. Если в одной хромосоме возникла полезная мутация, а в гомологичной хромосоме, пришедшей от другого родителя, также есть полезная мутация, но в другом локусе, то совместиться они могут только путем кроссинговера. Тогда такая рекомбинантная хромосома принесет популяции объединение двух полезных признаков [12].
Зависимость уровня рекомбинаций от среды можно рассматривать как оптимальную стратегию рекомбинационной системы, которая состоит в том, что достигается минимум рассеивания потенциальной генетической изменчивости, когда потребности в ней минимальны, т.е. в благоприятных условиях [13].
Линия yw была получена в результате следующих скрещиваний:
P: +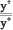 >
> 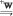

F1: + 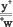 >
> 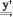
F2: ; 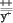 ; ;
; ; 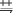 ;
;
; 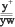 ; ;
; ; 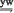
Из потомства F2 отбирали желтотелых белоглазых самцов () и скрещивали с желтотелыми красноглазыми самками ( и ). Самки в срещивании с самцами давали поколение линии yw.
P: + >
F3: ; 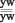 ; ;
; ;
Из поколения F3 (2-го скрещивания) отбирали желтотелых белоглазых самцов и самок (и ) и ставили их на массовое размножение.
Изучение частоты спонтанного кроссинговера между генами желтое тело (y) и белые газа (w), находящимися I хромосоме, в мы проводили по следующему скрещиванию:
P: + >
F1: + 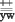 >
>
F2: ; ; ; ; ; ; ; 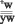
В ходе нашего исследования в потомстве велся учет мух всех фенотипических классов, как среди самок, так и среди самцов. Спонтанный кроссинговер определялся отдельно по мужским и женским особям, а также по всему потомству (таблица1).
Таблица 1
Уровень спонтанного кроссинговера у линии yw/++
|
№ культуры |
5 |
11 |
15 |
17 |
18 |
||||||||||
|
+ |
> |
- |
+ |
> |
- |
+ |
> |
- |
+ |
> |
- |
+ |
> |
- |
|
|
rf (y-w)(%) |
- |
2,7 |
1,04 |
- |
2,86 |
1,56 |
- |
10 |
3,5 |
- |
5,56 |
2,78 |
1,67 |
- |
0,93 |
|
2 (rfst=1,5%) |
- |
0,96 - |
0,14 - |
- |
1,23 - |
0,24 |
- |
48,17 +++ |
2,67 - |
- |
10.99 +++ |
1,09 - |
1,93 - |
- |
21,66 +++ |
Условные обозначения: «+++» - достоверные отличия при P<0,001 (χ2st=10,89) и k=1; «++» - при P<0,01 (χ2st =6,64) k=1; «+» - при P<0,05 (χ2st =3,84) k=1.
Из 18 пробирных культур только в 5 был зафиксирован кроссинговер, что составило 28%. Это возможно вследствие следующих причин: во-первых, из-за небольшой численности потомства, так как мутантная линия yw с двумя рецессивными мутациями обладает пониженной жизнеспособностью и, во-вторых, маленькое расстояние между генами y и w - 1,5% приводит к появлению небольшого числа рекомбинантного потомства.
Рассматривая кроссинговер у особей разного пола, отметим, что чаще он наблюдался в Fа среди самцов (4 случая), нежели у самок (1 случай). Скорее всего отрицательную роль в этом феномене у гетерозиготных самок +w/yw и w+/yw вновь играет хромосома - анализатор yw, которая увеличивает смертность у её носителей. Самцы - гемизиготный пол (XY) не имеют X хромосомы с генами yw, Y-хромосома «вредного влияния» на кроссоверных мух не оказывает. Этими же причинами можно объяснить статистически более значимую величину кроссинговера у самцов (rfср=4,59%, +), чем у самок (rfср=1,67%) - рекомбинантные особи женского пола чаще не выживают.
Если сравнивать частоту спонтанного кроссинговера у разных полов и величину общего кроссинговера со стандартным значением, определенным по генетической карте - 1,5%, мы выявляем следующее: у самцов она несколько выше, у самок находится в пределах нормы; среднее значение кроссинговера по всем культурам составляет 1,69, что статистически не отличается от табличного значения.
Рассматривая флуктуации показателей rf по повторностям (таблица 2) следует объяснить разной чувствительностью стадий оогенеза к внешним воздействиям [7] .
Таблица 2
Значение χ2 в каждой повторности на стандартной среде
|
№ повторности |
№ 1 |
№ 2 |
№3 |
№4 |
№5 |
|
rf |
3,19 |
1,83 |
0,33 |
2,057 |
0,00 |
|
χ2 (для rf=1,5%) |
25,96(+++) |
10,75(+) |
5,77(-) |
2,783(-) |
14,584(++) |
Условные обозначения: «+++» - достоверные отличия при P<0,001 (χф=16,27) и к=3; «++» - достоверные отличия при P<0,01 (χф=11,34) и к=3; «+» - достоверные отличия при P<0,5 (χф=7,82) и к=3; «-» - достоверных отличий нет.
Данные анализа, представленные в таблице 2, показывают, что в первой повторности отличия в высшей степени достоверны (χф=25,96 > χф=16,27), о чём свидетельствует высокое значение уровня рекомбинации rf (yw)=3,19% (табл. 2). Именно эта стадия на основе структурно – морфологических особенностей ядра в онтогенезе дрозофилы относится к ооцитам I класса клеток [7]. Данный класс включает клетки на синаптической стадии мейоза. Ядро таких клеток содержит синаптонемный комплекс – основной атрибут кроссинговера. Поэтому закономерно, что ооциты на этой стадии является рекомбиногенно чувствительным к любому внешнему и внутреннему воздействию. Следовательно, наиболее чувствительным этапом развития ооцитов является ранняя стадия их созревания.
В ходе эксперимента было поставлено 30 индивидуальных культур yw/++ × yw/Y. Потомство дали 18 родительских пар, 12 культуры не развились вследствие стерильности одного или двух родителей (40%). На жизнеспособных мух могли повлиять мутации y и w, которые значительно снижают численность гомозиготных по этим генам самок, по сравнению с гетерозиготными особями. Из 18 культур в 13 случаях статистически достоверно преобладали мухи дикого типа, в 3 вариантах особи генотипа yw вовсе отсутствовали.
В нашем эксперименте вторичная половая пропорция (>/+) в 11 культурах была меньше 1, в 5 культурах - больше 1 и в 2 культурах равна 1. Общая численность самок - 668, самцов - 515, разница статистически достоверна (χ2=19,8 +++ при P>0,001,k=1). Такая диспропорция в сторону женского пола может свидетельствовать об относительно стабильных условиях существования популяции. Кроме того, согласно концепции В.А. Геодакяна, у самцов одна X-хромосома, а у самок 2 X-хромосомы, одна из которых может компенсировать поврежденный локус в гомологичной половой хромосоме, поэтому самки живут дольше и у них норма реакции шире, чем у самцов [2].
Проведённое исследование показало возможность использования линии Drosophila melanogaster yw в решении частных вопросов генетики и экологии.
Библиографическая ссылка
Жестков В.А., Сидорская В.А. СОЗДАНИЕ ЛИНИИ DROSOPHILA MELANOGASTER CO СЦЕПЛЕННЫМИ МАРКЕРАМИ YW И ИЗУЧЕНИЕ ЕЁ БИОЛОГИЧЕСКИХ ОСОБЕННОСТЕЙ // Научное обозрение. Биологические науки. 2017. № 1. С. 91-94;URL: https://science-biology.ru/ru/article/view?id=1038 (дата обращения: 03.07.2026).