

 science-review.ru
science-review.ru
Теоретические исследования в поиске прогрессивных знаний в области изучения генома сельскохозяйственных кур позволяют разрабатывать и усовершенствовать программы по селекции и разведению кур, нацеленные на повышение продуктивных качеств с одновременным улучшением устойчивости к стрессам и болезням.
Цель исследования – изучение эволюционного развития и молекулярно-генетического потенциала генома сельскохозяйственных кур.
Задачи исследования: изучение гипотез о происхождении сельскохозяйственных кур; раскрытие ключевых особенностей, характерных для кариотипа и генома сельскохозяйственных кур; анализ особенностей эволюции генома сельскохозяйственных кур.
Материалы и методы исследования
Работа выполнена на основе теоретических исследований. С марта по ноябрь 2023 года проводились поиск и анализ научной литературы по исследуемой теме. Всего для работы над статьей на цифровых отечественных и международных платформах (Web of Science, Scopus, Научная электронная библиотека и др.) изучено и проанализировано за период с 1868 по 2023 год более 580 научных трудов, из которых 48 были использованы в нашей работе.
Результаты исследования и их обсуждение
Класс птиц расположен на филогенетическом древе в эволюционном промежутке между пресмыкающимися и млекопитающими [1] и насчитывает более 10 тыс. видов, но к сельскохозяйственным относят только 8 [2]. Среди сельскохозяйственных птиц особое место занимают куры, которые принадлежат к отряду Galliformes (Курообразные), семейству Phasianidae (Фазановые), роду Gallus (Джунглевые куры; отличаются наличием гребня и сережек), виду Gallus gallus (Банкивская джунглевая курица) [2].
Сельскохозяйственное куроводство обеспечивает рынок диетическими продуктами питания – мясом и яйцами [3]. Ежегодно в аграрном секторе экономики для производства мяса выращивается более 71 млрд птиц [4]. В мире производится мяса бройлеров более 103,4 млн т [5], яиц – более 80 млн т [6].
Согласно данным Продовольственной и сельскохозяйственной организации (FAO), мировая популяция кур составляет более 34,4 млрд голов, из которых больше половины (около 56%) приходится на Азию. В мире насчитывается более 1 600 пород кур. Куры составляют около 92% мирового поголовья домашней птицы, за ними следуют утки (5%) и индейки (2%). Куроводство, таким образом, обеспечивает мировое производство мяса птицы на 89% и яиц на 92% [4].
В Казахстане численность поголовья сельскохозяйственных птиц на 01.01.2023 г. составила 49,9 млн голов [7], из которых на конец 2021 г. почти 72,2% сосредоточено в пяти областях Казахстана – Алматинской 25,7%, Акмолинской – 19,7%, Костанайской – 10,1%, Северо-Казахстанской – 8,4%, Восточной-Казахстанской – 8,3% [8]. Производство же яиц в Казахстане на 01.01.2023 г. составило 5052,2 млн шт., при этом на долю куриных приходится 99,5% [7].
Следует отметить, что потребление мяса на душу населения из года в год, согласно данным ФАО, увеличивается, на первом месте стоит потребление мяса кур [4]. По оценкам, мясо кур удовлетворяет более 30% всех потребностей в животном белке [9].
Устойчивое развитие промышленного птицеводства (бройлерного, яичного) основано на интенсивном производстве [10], где поголовье варьирует от нескольких тысяч до нескольких миллионов голов, содержится в закрытых помещениях (напольное, клеточное содержание) с автоматическими системами кормления и поения [11], с очень высокой плотностью (33 кг м-2 или выше) [12]. Наличие кормов (зерно, соя и семена масличных культур) часто зависит от внешних производителей [11]. Использование воды (как в производстве кормов, так и в птицеводстве) является значительным [13] и составляет при производстве куриного мяса и яиц 4325 и 3265 м3 т-1 соответственно [14].
Увеличение производства продукции куроводства как одной из скороспелой отрасли животноводства обеспечивается путем усовершенствования генетических хозяйственно полезных качеств кур, улучшения условий их кормления и содержания [15]. Поэтому изучение генетики сельскохозяйственных кур (лат. Gallus domesticus) актуально и имеет большое значение для устойчивого улучшения программ селекции и разведения кур [16].
Контролируемая человеком искусственная эволюция сельскохозяйственных кур в условиях промышленного куроводства направлена [2] на улучшение резистентности и на повышение общей изменчивости кур (особенно массы и формы тела), повышение продуктивности и плодовитости кур при одновременном снижении затрат кормов на производимую куроводческую продукцию, приспособляемость кур к условиям промышленного содержания за счет изменений в физиологии и в поведении кур, в последнем случае – за счет снижения их пугливости при контакте с человеком.
Происхождение и одомашнивание сельскохозяйственных кур
Вид сельскохозяйственных кур (Gallus gallus domesticus) является современным потомком динозавров и первым амниотом, не относящимся к млекопитающим [17]. Современные птицы (Ornithurae) произошли от динозавров-терапод [18] в середине мезозойской эры [19].
Как видно из рисунка 1, Археоптерикс (обозначен звездочкой) считается первой известной птицей, жившей примерно 150 млн лет назад [19]. Эволюционное развитие генома курицы протекает отдельно от геномов млекопитающих и длится приблизительно в течение 310 млн лет [19].
Происхождение домашней курицы, в том числе и с точки зрения географического обитания, было предметом споров с тех пор, как Дарвин впервые предположил теорию монофилетического происхождения кур. Согласно теории Дарвина, индийская красная джунглиевая курица (Gallus gallus) является единственным предком всех домашних кур [20].
В основе теории Дарвина лежали следующие ключевые положения [20]: наличие заметного морфологического сходства между домашней курицей и красной джунглевой птицей; скрещивание этих двух видов дало плодородное потомство, тогда как скрещивание домашних кур с тремя другими видами джунглевых птиц приводило к низкой выживаемости цыплят.
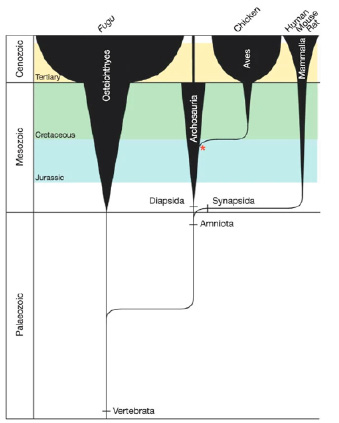
Рис. 1. Эволюция позвоночных с указанием существующих видов, чьи геномы были секвенированы [19]
Альтернативную гипотезу происхождения домашних кур – многовидовое, или полифилетическое, происхождение – впервые предложил Ф. Хатт в своей книге «Генетика домашней птицы» (1949). Он утверждал, что потенциально все четыре диких вида джунглевых птиц могли внести определенный вклад в генетический состав домашней курицы. Свою гипотезу он основывал на том факте, что ряд фенотипов, наблюдаемых у домашних кур, не обнаруживается в популяции красных джунглевых птиц, но присутствует у других видов джунглевых птиц [21].
Одомашнивание кур произошло в Азии примерно к 5400 г. до н.э. или уже к 8000 г. до н.э. [22]. Предположение Дарвина о том, что красная джунглевая курица была ближайшим предком домашней курицы, подтвердилось позже при анализе митохондриальной ДНК. Следовательно, красная джунглевая курица является признанным предком домашней курицы по материнской линии [23].
С момента одомашнивания красной джунглевой птицы (Gallus gallus) (примерно с 8000 по 5400 год до н.э.) в Азии [24] домашние куры (Gallus Gallus domesticus) подвергались комбинированному воздействию естественного и искусственного отбора. Это привело к заметному генетическому разнообразию по ряду признаков [25]. На современном этапе предполагается, что современная курица (Gallus gallus domesticus) была одомашнена от красной джунглевой курицы (RJF) [20], но с некоторым участием еще одного близкородственного вида – серой джунглевой курицы [24]. Также предполагается, что азиатская красная джунглевая курица (Gallus gallus) (RJF), обитающая на юго-востоке Азии, является родоначальником всех домашних кур, которая была одомашнена в долине Инда еще в 3200 г. до н.э. [26]. Более поздние археологические раскопки в Китае указывают на наличие более ранних центров приручения [24].
G. gallus, дикий RJF, обитающий в лесах Юго-Восточной Азии и Индии, распространился после одомашнивания в процессе переселения людей в другие части мира, что привело к появлению многих пород кур [27].
Домашние куры принадлежит к роду Gallus, который таксономически включает четыре диких вида джунглевых птиц: красную джунглевую (Gallus gallus; RJF), серую джунглирующую (G. sonneratii), цейлонскую джунглевую (G. lafayetii) и зеленую джунглевую (G. varius). Все четыре вида джунглевых птиц обитают в разных географических регионах Южной Азии и имеют довольно большие различия в морфологии. В настоящее время существует 5 подвидов RJF: G. g. gallus (Юго-Восточная Азия RJF) и G. g. spadiceus, G. g. bankiva, G. g. murghi (индийский RJF) и G. g. Jabouillei. Эти классификации, в основном, базируются на фенотипических признаках и географическом распределении популяций [28]. В настоящее время на существование красной джунглевой птицы (RJF) все большее влияние оказывают разрушение среды обитания, незаконная охота и потребление хищниками, что может в конечном итоге привести к ее исчезновению [29].
Таким образом, долгая история селекции и исследований сельскохозяйственных кур создала богатую коллекцию генотипического и фенотипического разнообразия в виде пород, линий и семейств со специфическими, продуктивными характеристиками.
В течение прошлого века домашняя курица превратилась в основной источник пищи за счет адаптации генома к специализированным яйцекладкам (несушкам) и быстрорастущим мясным птицам (бройлерам), продуктивность которых намного превосходит продуктивность их диких предков. Большинство современных сельскохозяйственных несушек производят примерно 300 яиц в год, в то время как красная джунглевая курица (RJF) обычно откладывает одну кладку из 5–9 яиц в год. Современные бройлеры быстро достигают массы тела 4–5 кг, а вес взрослого самца красной джунглевой птицы (RJF) – около 1 кг [27]. Коммерческие поставщики бройлеров и несушек производят более 71 млрд голов птицы в год, чтобы удовлетворить текущие мировые потребности потребителей в более 104 млн т мяса и более 1,3 трлн шт. яиц [5]. Как видим, искусственный отбор в процессе одомашнивания кур и последующая специализация на курах мясного (бройлеры) и яичного (несушки) типа внесли в молекулярную организацию генома современных сельскохозяйственных кур существенные эволюционные коррективы [30].
Кариотип сельскохозяйственных кур
Кариотип как физическая и генетическая карта генома с низким разрешением строится с учетом системного распределения хромосом (идеограмма). «Хромосомология» вида – это первый шаг в картировании генома. Хромосомы кур изучаются с применением двух методов – окрашивания и картирования известных клонов и использования их в качестве ориентиров для хромосом. Кариотипы птиц существенно отличаются от кариотипов млекопитающих.
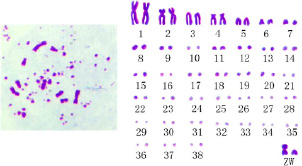
Рис. 2. Кариотип сельскохозяйственных кур (Gallus gallus domesticus) [19]
Большинство кариотипов птиц, в том числе и кур, содержат хромосомы, которые заметно различаются по размерам, поэтому их называю макро- и микрохромосомами [31].
Ключевые особенности, характерные для кариотипа сельскохозяйственных кур, следующие.
1. Количество хромосом у кур в составе кариотипа велико:
– диплоидный набор хромосом равен 78 (2n = 78), гаплоидный – 39 (n = 39) [30, 31].
2. Кариотип домашних кур устроен сложно [32–34], о чем свидетельствует рисунок 2.
3. Диплоидный набор сельскохозяйственных кур, состоящий из 39 пар хромосом (2n = 78), включает 38 пар аутосом и 1 пару половых хромосом. Половые хромосомы обозначены буквами Z и W (рис. 2), причем гетерогаметным полом является курица (♀ ZW), гомогаметным – петух (♂ ZZ) [19].
4. Разброс в размерах хромосом – это цитогенетическая особенность кариотипа домашних кур [30].
5. Из 38 пар аутосом к крупным относят 5 хромосом (макрохромосомы), к промежуточным – 5 и к малым по размеру – 28 хромосом (микрохромосомы) [30].
В работе J. Masabanda и M. Schmid отмечено, что из 39 пар хромосом к крупным аутосомам – макрохромосомам – отнесены 10 пар (хромосомы 1–10), к микрохромосомам – 28 пар (хромосомы 11–38), а пара половых хромосом – W и Z – представлены как макрохромосома (хромосома Z) и микрохромосома (хромосома W), последняя из которых имеется у гетерогаметного пола (♀ ZW) [30, 35]. Согласно K. Ladjali и D.W. Burt, 39 пар хромосом включают девять пар (вместе с половыми хромосомами Z и W), цитологически различные макрохромосомы и 30 микрохромосом [10].
Из вышеизложенного можно заключить, что способность различать хромосомы кур является одним из приоритетов в селекционно-генетической работе.
Геном сельскохозяйственных кур и особенности его организации
Гаплоидное содержание генетического аппарата в геноме кур равно 1,2 × 109 пар оснований ДНК [36] и разделено, как это было отмечено выше, между 39 хромосомами.
На сегодня геном кур изучен на 96% (1,03 ГБ) от его прогнозируемого размера, поскольку раскрыты последовательности для всех 10 крупных аутосом, 19 из 28 микрохромосом и обеих половых хромосом [35].
Генетическая карта генома сельскохозяйственных кур включает около 650 молекулярных маркеров [37]. Генетический размер равен 2500–3400 сМ [38].
Средний размер макрохромосом составляет примерно 130 Мб. К микрохромосомам относят хромосомы, размеры которых варьируют в пределах от 0,5 до 1,5 мкм. Средний размер микрохромосом составляет 12,5 Мб, причем самый маленький из них равен 7 Мб. На микрохромосомы приходится 30% генома сельскохозяйственных кур [33].
Тридцать микрохромосом кур содержат около одной трети геномной ДНК, т.е. в них содержится примерно в два раза больше генов, чем в макрохромосомах. Микрохромосомы представляют собой богатую генами ДНК, и в них сосредоточено, по крайней мере, 50% генов. Микрохромосомы демонстрируют высокую скорость рекомбинации, они реплицируются в течение первой половины S-фазы [39].
По сравнению с макрохромосомами большинство микрохромосом имеют более низкое содержание AT-пар (A + T) и более высокое – GC-пар (G + C) [40].
Особенности генома сельскохозяйственных кур:
1. Общий размер генома кур по сравнению с геномом млекопитающих небольшой и составляет 1,1 Гб [40], тогда как, например, у мыши он равен 2,5 Гб [41], у человека – 2,9 Гб [42].
2. Хромосомы кур значительно различаются по размеру, поэтому их классифицируют на микро- и макрохромосомы. Согласно соглашению Международного консорциума по секвенированию генома курицы (ICGSC 2004 [19, 43]), хромосомы кур классифицируют на три класса:
– пять макрохромосом (GGA 1-5), в среднем длиной от ~ 50 до 200 Мб;
– пять промежуточных хромосом (GGA 6-10), в среднем длиной от 20 до 40 Мб;
– 28 микрохромосом (GGA 11-38), в среднем длиной примерно 12 Мб.
3. Микрохромосомы кур от общего генома занимают только 18%, но содержат примерно 31% всех генов (ICGSC 2004 [19]), что указывает на плотную генную структуру с короткими межгенными последовательностями, которые в 3–4 раза короче по сравнению с генами макрохромосом [43].
4. Содержание GC выше на микрохромосомах, как и уровень метилирования, за счет увеличения количества сайтов CpG [40].
5. Микрохромосомы реплицируются раньше, чем макрохромосомы [19]. Скорость рекомбинации выше на микрохромосомах (средняя скорость 6,4 сМ/Мб), чем на промежуточных хромосомах (3,9 сМ/Мб) и на макрохромосомах (2,8 сМ/Мб) [19, 43].
6. Микрохромосомы богаты генами, поскольку содержат в два раза больше генов, чем макрохромосомы [19].
7. Карта генетического сцепления кур, созданная для исследования еще в начале прошлого века, в настоящее время включает 2172 генетических локусов с общей длиной около 4000 сМ [44].
7.1. Геном кур с 2400 генами расшифрован в 2004 г. На сегодня исследовано примерно 1000 генов [19, 45].
7.2. Геном кур состоит примерно из одного миллиарда пар ДНК-последователь-ностей и включает примерно 20 000–23 000 генов [19].
8. Большинство микрохромосом являются акроцентрическими [40].
Особенности эволюции генома сельскохозяйственных кур
Курица имеет общего предка с млекопитающими около 300–350 миллионов лет назад и считается в исследованиях эволюционного процесса ключевым видом. Изучение генома животных в сравнении с геномом кур используется для изучения эволюции конкретных генов или семейств генов либо самой организации генома [23].
Производственно-ориентированный отбор в куроводстве способствовал порождению основных генетических изменений, произошедших у современных сельскохозяйственных кур в процессе эволюции [30].
Эволюционный процесс генома сельскохозяйственных кур описывают следующими ключевыми особенностями [19]:
– геном курицы в 3 раза меньше генома млекопитающих за счет существенного уменьшения содержания повторов, псевдогенов и сегментарных дупликаций;
– низкая скорость транслокаций хромосом в обеих линиях (курица, человек) от последнего общего предка, тогда как внутрихромосомные перестройки (например, инверсии) встречаются чаще;
– расширение и сокращение мультигенных семейств является, по предположению, основным фактором независимой эволюции млекопитающих и птиц;
– скорость синонимичных замен в геноме кур повышена для генов как в микрохромосомах, так и в субтеломерных областях макрохромосом;
– геном курицы обладает высокой специфичностью обратной транскриптазы от LINE-последовательности;
– в отличие от геномов всех других позвоночных, изученных до сих пор, в геноме курицы не было активных SINEs элементов в течение последних приблизительно 50 млн лет;
– в геноме кур идентифицировано примерно 70 Мб последовательности, которая, с большой вероятностью, функционирует и в геноме человека;
– в геноме птиц примерно 274 генов, которые содержатся в геноме человека, отсутствуют [46].
В мире существует множество фенотипически различимых пород и линий домашних кур, что предоставляет уникальную научно-практическую возможность для изучения истории формирования современных сельскохозяйственных кур и определения признаков отбора, являющегося результатом как процесса одомашнивания, так и воздействия окружающей среды и технологии ведения куроводства на промышленном уровне [47, 48].
Тенденция утраты генетического биоразнообразия кур отражена в глобальных планах действий ФАО [47].
Куры яичных и мясных пород и линий в промышленном куроводстве становятся все менее разнообразными. Ключевым фактором, влияющим на снижение генетической изменчивости в промышленном куроводстве, является резкое снижение числа пород кур, используемых в селекционных программах. Так, в состав современных промышленных кроссов входят примерно 4–7 (до 10) пород из более чем 1600 известных во всем мире пород, притом что из 1600 известных во всем мире пород 2% уже исчезли, 30% грозит исчезновение, 63% подлежат сохранению, 26% находятся вне зоны риска, а для 41% с тепень риска неизвестна [2].
Заключение
Изучение вопроса происхождения и одомашнивания сельскохозяйственных кур показало, что данный вопрос все еще остается предметом споров, так как до настоящего времени каждая из двух теорий актуальна в научных работах: согласно монофилетической теории (Ч. Дарвин, 1868 г.), индийская красная джунглевая курица является единственным предком для всех домашних кур, а по альтернативной полифилетической гипотезе (Ф. Хатт, 1949 г.) все четыре диких вида джунглевых птиц могли внести определенный вклад в генетический состав домашней курицы. Функциональная аннотация геномов сельскохозяйственных птиц имеет решающее значение для понимания молекулярных механизмов экономически важных и биологически и селекционно сложных признаков, таких как рост, размножение, продуктивность и устойчивость к инфекционным болезням.
При изучении вопросов о происхождении и одомашнивании сельскохозяйственных кур определено, что:
– вид сельскохозяйственных кур (Gallus gallus domesticus) является современным потомком динозавров и первым амниотом, не относящимся к млекопитающим;
– одомашнивание кур произошло в Азии примерно к 5400 г. до н.э. или уже к 8000 г. до н.э.;
– современная курица была одомашнена от красной джунглевой курицы, но с некоторым участием еще одного близкородственного вида – серой джунглевой курицы.
Изучение особенностей кариотипа, а также организации и эволюции генома сельскохозяйственных кур показало, что:
– разброс в размерах хромосом – это цитогенетическая особенность кариотипа домашних кур;
– гаплоидное содержание генетического аппарата в геноме кур равно 1,2 × 109 пар оснований ДНК и разделено между 39 хромосомами;
– хромосомы кур классифицируют на три класса: пять макрохромосом, в среднем длиной от ~ 50 до 200 Мб; пять промежуточных хромосом, в среднем длиной от 20 до 40 Мб, и 28 микрохромосом, в среднем длиной примерно 12 Мб;
– микрохромосомы богаты генами (содержат в два раза больше генов, чем макрохромосомы);
– в результате обширных программ разведения было выведено около 60 пород кур, представляющих четыре различные линии: яичный тип, дичь, мясной тип и бантам (декоративная карликовая порода);
– курица имеет общего предка с млекопитающими около 300–350 млн лет назад;
– ключевым фактором, влияющим на снижение генетической изменчивости в промышленном куроводстве, является резкое снижение числа пород кур, используемых в селекционных программах.
Библиографическая ссылка
Мельников Н.П., Багыбаева А.М., Исагулов Т.Е., Мельникова Т.В., Джарбанова А.Д., Булах А.В., Безрукова А.Н., Мусина Г.Ш., Джамалова Г.А. Происхождение, генетика и особенности эволюции генома сельскохозяйственных кур // Научное обозрение. Биологические науки. 2024. № 1. С. 71-78;URL: https://science-biology.ru/ru/article/view?id=1357 (дата обращения: 03.07.2026).
DOI: https://doi.org/10.17513/srbs.1357